Therapeutischer Manuka-Honig: Nicht mehr so alternativ

Die Forschung zu medizinischem Honig erlebt eine bedeutende Renaissance. Von einem volkstümlichen Heilmittel, das von der Schulmedizin weitgehend als „alternativ“ abgetan wurde, zeigt sich nun ein wachsendes Interesse von Wissenschaftlern, klinischen Praktikern und der breiten Öffentlichkeit an den therapeutischen Anwendungen von Honig. Dieses Interesse wird durch mehrere Faktoren angetrieben: Erstens hat der Anstieg der Antibiotikaresistenz vieler bakterieller Krankheitserreger das Interesse an der Entwicklung und Nutzung neuartiger antibakterieller Mittel geweckt; zweitens haben eine zunehmende Anzahl zuverlässiger Studien und Fallberichte gezeigt, dass bestimmte Honige sehr wirksame Wundbehandlungen sind; drittens erzielt therapeutischer Honig einen Premiumpreis, und die Honigindustrie fördert aktiv Studien, die es ihr ermöglichen, davon zu profitieren; und schließlich bietet die sehr komplexe und recht unvorhersehbare Natur von Honig eine attraktive Herausforderung für Laborswissenschaftler. In diesem Artikel geben wir einen Überblick über die Manuka-Honig-Forschung, von Beobachtungsstudien zu seinen antimikrobiellen Effekten bis hin zu aktuellen experimentellen und mechanistischen Arbeiten, die darauf abzielen, Honig in die Schulmedizin zu integrieren. Wir skizzieren aktuelle Wissenslücken und verbleibende Kontroversen darüber, wie Honig wirkt, und schlagen neue Studien vor, die Honig zu einer nicht mehr „alternativen“ Alternative machen könnten.
Einleitung
Honig wurde im Laufe der Geschichte der Menschheit als Medizin verwendet. Eine der häufigsten und beständigsten therapeutischen Anwendungen von Honig war die Verwendung als Wundauflage, fast sicher aufgrund seiner antimikrobiellen Eigenschaften. Mit dem Aufkommen hochwirksamer Antibiotika in den 1960er Jahren wurde Honig als „wertlos, aber harmlos“ abgetan (Soffer, 1976). Die aktuelle und wachsende Krise der Antibiotikaresistenz hat jedoch das Interesse an der Verwendung von Honig wiederbelebt, sowohl als wirksames Mittel an sich als auch als therapeutische Grundlage zur Entwicklung neuer Behandlungsmethoden. Honig wird üblicherweise aus dem Nektar von Blüten gewonnen und von Bienen produziert, meist von der Europäischen Honigbiene Apis mellifera, und ist eine komplexe Mischung aus Zuckern, Aminosäuren, Phenolen und anderen Substanzen. Honigsorten, die von verschiedenen Blütenpflanzen stammen, unterscheiden sich erheblich in ihrer Fähigkeit, Bakterien abzutöten, was die Literatur über Honig verkompliziert und es manchmal schwierig macht, Ergebnisse zwischen verschiedenen Studien zu reproduzieren (Allen et al., 1991; Irish et al., 2011). Die Mehrheit der aktuellen Studien, die den Wirkmechanismus von Honig untersuchen, konzentriert sich auf gut charakterisierten, standardisierten aktiven Manuka-Honig, der von bestimmten Leptospermum-Arten stammt, die in Neuseeland und Australien heimisch sind und als Wundpflegeprodukt bei den entsprechenden medizinischen Zulassungsbehörden registriert sind. Sofern nicht anders angegeben, wird sich diese Übersicht auf Manuka-Honig konzentrieren.
Chemische Analysen von aktivem Manuka-Honig
Professor Peter Molan von der Universität Waikato in Neuseeland war der erste, der die ungewöhnliche Aktivität von Manuka-Honig berichtete und Mitte der 1980er Jahre begann, seine Wirkung gegen eine Vielzahl verschiedener Bakterienarten zu testen. Es war zwar klar, dass bereits niedrige Konzentrationen von Manuka-Honig bakterielle Krankheitserreger abtöten, doch die spezifische aktive Substanz, die dafür verantwortlich ist, blieb viele Jahre lang unbekannt. Hoher Zuckergehalt und niedriger pH-Wert machen Honig hemmend für das mikrobielle Wachstum, aber die Aktivität bleibt erhalten, wenn diese auf vernachlässigbare Werte verdünnt werden. Viele verschiedene Honigsorten produzieren ebenfalls Wasserstoffperoxid, wenn Glukoseoxidase, die von der Honigbiene stammt, mit Glukose und Wasser reagiert. Im Manuka-Honig ist die Wasserstoffperoxidproduktion jedoch relativ gering und kann durch Katalase neutralisiert werden, dennoch bleibt die Aktivität erhalten. Die Ursache dieser verbleibenden Aktivität, die als „Nicht-Peroxid-Aktivität“ oder NPA bezeichnet wird, wurde schließlich 2008 enthüllt, als zwei Labore unabhängig voneinander Methylglyoxal (MGO) im Manuka-Honig identifizierten (Adams et al., 2008; Mavric et al., 2008). MGO entsteht durch die spontane Dehydratisierung seines Vorläufers Dihydroxyaceton (DHA), einem natürlich vorkommenden Pflanzenstoff, der im Nektar der Blüten von Leptospermum scoparium, Leptospermum polygalifolium und einigen verwandten Leptospermum-Arten heimisch in Neuseeland und Australien vorkommt (Adams et al., 2009; Williams et al., 2014; Norton et al., 2015). MGO kann relativ unspezifisch mit Makromolekülen wie DNA, RNA und Proteinen reagieren (Adams et al., 2008; Mavric et al., 2008; Majtan et al., 2014b) und könnte theoretisch für Säugetierzellen toxisch sein (Kalapos, 2008). Es gibt jedoch keine Hinweise auf Schäden an Wirtszellen, wenn Manuka-Honig entweder oral konsumiert oder als Wundauflage verwendet wird; tatsächlich scheint Honig die Heilung zu fördern und Narbenbildung zu reduzieren, wenn er auf Wunden aufgetragen wird (Biglari et al., 2013; Majtan, 2014; Dart et al., 2015). Wie es diese scheinbar selektive Toxizität gegenüber bakteriellen Zellen ausübt, ist nicht bekannt.
Hohe Konzentrationen von MGO oder Wasserstoffperoxid führen in der Regel zu dem aktivsten Honig, jedoch ist die Korrelation nicht immer perfekt, was darauf hindeutet, dass andere Bestandteile des Honigs die Aktivität modulieren können (Molan, 2008; Kwakman et al., 2011; Chen et al., 2012; Lu et al., 2013). Bee defensin-1, ein antimikrobielles, von Bienen stammendes Peptid, ist für die Aktivität im Revamil-Honig verantwortlich, einem aktiven Honig aus einer nicht offengelegten Quelle, scheint jedoch strukturell modifiziert und in Manuka-Honig inaktiv zu sein (Kwakman et al., 2011; Majtan et al., 2012). Der Gehalt an Leptosin, einem ausschließlich in Leptospermum-Honig vorkommenden Glycosid, korreliert mit der Wirksamkeit und könnte die antimikrobielle Aktivität von Manuka-Honig modulieren (Kato et al., 2012). Ebenso können verschiedene phenolische Verbindungen mit potenzieller antimikrobieller Wirkung vorhanden sein, insbesondere in dunkler gefärbten Honigen, und obwohl diese in Konzentrationen vorkommen, die allein wahrscheinlich nicht hemmend wirken, können sie synergistisch miteinander oder mit anderen Honigbestandteilen wirken, um die Aktivität zu erzeugen oder zu verändern (Estevinho et al., 2008; Stephens et al., 2010). Phenole können auch als Antioxidantien wirken und könnten für die entzündungshemmenden und wundheilenden Eigenschaften von Honig verantwortlich sein (Stephens et al., 2010). Es sollte beachtet werden, dass nicht alle Leptospermum-Arten aktiven Honig produzieren, und selbst innerhalb von L. scoparium und L. polygalifolium-Honig können die MGO-Werte von etwa 100 bis über 1200 ppm reichen (Windsor et al., 2012). Eine Untersuchung der Aktivität australischer Honige ergab, dass Honig von Leptospermum-Pflanzen, die an der Grenze zwischen New South Wales und Queensland wachsen, besonders aktiv war, wobei jedoch nicht bekannt ist, ob dies auf Pflanze, Boden, Klima oder andere Faktoren zurückzuführen ist (Irish et al., 2011).
Die Hemmung von Krankheitserregern durch Honig
Honig wurde in vitro an einer Vielzahl von Krankheitserregern getestet, insbesondere solchen, die Haut, Wunden und Schleimhäute besiedeln können, wo eine topische Honiganwendung möglich ist. Bis heute haben in vitro-Tests gezeigt, dass Manuka-Honig alle getesteten problematischen bakteriellen Krankheitserreger wirksam hemmen kann (zusammengefasst in Tabelle 1). Besonders interessant ist, dass klinische Isolate mit multiplen Resistenzen (MDR) keine verringerte Empfindlichkeit gegenüber Honig zeigen, was auf ein breites Wirkungsspektrum hinweist, das kein bekanntes antimikrobielles Mittel aufweist (Willix et al., 1992; Blair und Carter, 2005; George und Cutting, 2007; Tan et al., 2009). Zudem waren Versuche, im Labor honigresistente Stämme zu erzeugen, nicht erfolgreich, und es gibt keine Berichte über klinische Isolate mit erworbener Honigresistenz (Blair et al., 2009; Cooper et al., 2010).
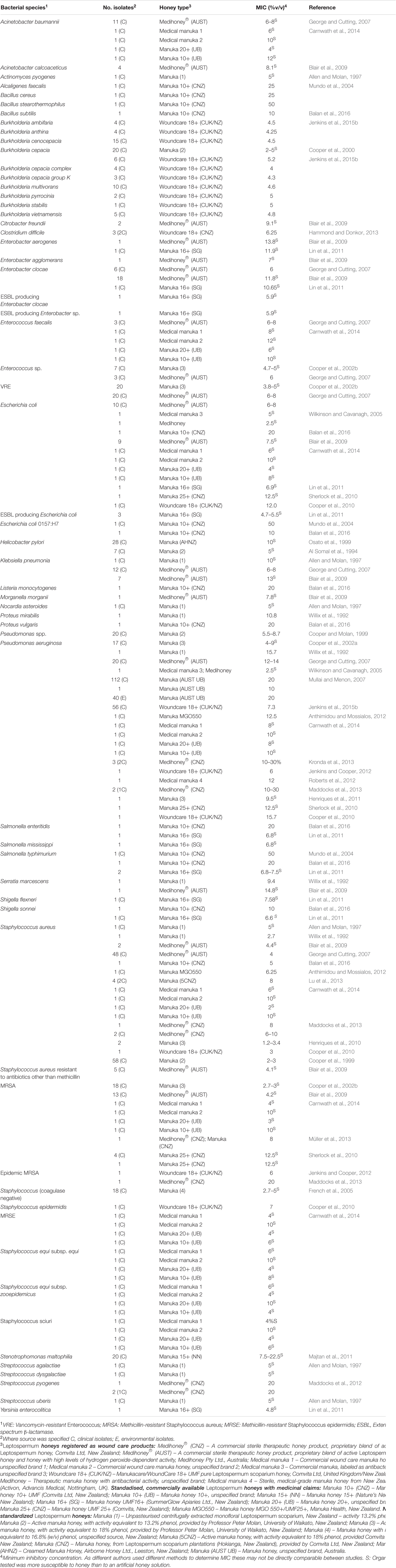
TABELLE 1. Bakterielle Arten, die als empfindlich gegenüber therapeutischem Manuka-Honig befunden wurden.
Neben der Hemmung planktonischer Zellen kann Honig auch Bakterien, die in Biofilmen leben, zerstreuen und abtöten. Biofilme sind Zellgemeinschaften, die in der Regel in einer selbstproduzierten extrazellulären Matrix eingeschlossen sind und an Oberflächen haften, einschließlich Wunden, Zähnen, Schleimhäuten und implantierten Geräten. Mikroben, die in Biofilmen leben, sind vor antimikrobiellen Mitteln geschützt und können anhaltende, nicht abklingende Infektionen verursachen. Manuka-Honig zerstört Zellaggregate (Maddocks et al., 2012; Roberts et al., 2012) und verhindert die Bildung von Biofilmen durch eine Vielzahl problematischer Krankheitserreger, darunter Streptococcus- und Staphylococcus-Arten, Pseudomonas aeruginosa, Escherichia coli, Proteus mirabilis, Enterobacter cloacae, Acinetobacter baumannii und Klebsiella pneumonia (Maddocks et al., 2012, 2013; Lu et al., 2014; Majtan et al., 2014a; Halstead et al., 2016). Wichtig ist, dass Honig auch etablierte Biofilme zerstören und die darin lebenden Zellen abtöten kann, obwohl dafür eine höhere Konzentration als bei planktonischen Zellen erforderlich ist (Okhiria et al., 2009; Maddocks et al., 2013; Lu et al., 2014; Majtan et al., 2014a). Ganz aktuell wurde Manuka-Honig an einem Mehrspezies-Biofilm getestet, der Staphylococcus aureus, Streptococcus agalactiae, Pseudomonas aeruginosa und Enterococcus faecalis enthielt, und es wurde festgestellt, dass die Lebensfähigkeit aller Arten außer E. faecalis reduziert wurde, das nicht eradiziert werden konnte (Sojka et al., 2016). Dies hat klare klinische Implikationen für die Anwendung von Honig bei Wunden mit Biofilmen, und das Verständnis, wie der Biofilm E. faecalis das Überleben ermöglicht, obwohl es normalerweise durch Honig abgetötet wird, ist ein wichtiges und interessantes zukünftiges Forschungsfeld. MGO scheint größtenteils, aber nicht vollständig für die Hemmung von Biofilmen durch Manuka-Honig verantwortlich zu sein, was erneut die Bedeutung zusätzlicher Komponenten hervorhebt, die die Aktivität modulieren (Kilty et al., 2011; Lu et al., 2014).
Das Wirkungsspektrum von Honig gegenüber nicht-bakteriellen Krankheitserregern ist noch nicht gut etabliert. Neuere Studien, die die antivirale Wirkung von Manuka-Honig untersuchten, legen nahe, dass er Potenzial für die Behandlung des Varizella-Zoster-Virus (Ursache von Windpocken und Gürtelrose) (Shahzad und Cohrs, 2012) und Influenza (Watanabe et al., 2014) besitzt. Pilzpathogene der Haut, darunter Candida albicans und Dermatophytenarten, sind gegenüber Manuka-Honig deutlich weniger empfindlich als Bakterien, werden jedoch durch Honig mit hohem Wasserstoffperoxid-Gehalt gehemmt (Brady et al., 1996; Irish et al., 2006). Manuka- und Nicht-Manuka-Honig konnten die Lebensfähigkeit von Sporen des Mikrosporidiers Nosema apis, einem wichtigen Bienenpathogen, reduzieren, konnten eine bereits bestehende Bieneninfektion jedoch nicht heilen (Malone et al., 2001). Es gibt nur sehr wenige Studien zur Anwendung von Honig bei Protozoen- oder Helminthenparasiten, und diese verwendeten keinen Honig mit gut charakterisierter Aktivität, was die Bewertung der Bedeutung ihrer Ergebnisse erschwert (Bassam et al., 1997; Nilforoushzadeh et al., 2007; Sajid und Azim, 2012).
Honig in der Schulmedizin: Neuere experimentelle und mechanistische Studien beleuchten, wie Honig wirkt
Aktiver Manuka-Honig ist weit verbreitet als therapeutisches Mittel und funktionelles Lebensmittel erhältlich, und die meisten Verbraucher akzeptieren ihn als ein ganzheitliches, etwas mysteriöses Produkt. Ein mangelndes Verständnis darüber, wie Honig Bakterien abtötet und die Heilung fördert, begrenzt jedoch seine Akzeptanz in der Schulmedizin, wo er noch als „alternativ“ oder „komplementär“ gilt. Die überwiegende Mehrheit der bisherigen Forschungsarbeiten zu Honig war beschreibend, doch neuere Studien versuchen, die Wirkungsweise von Honig zu entschlüsseln und verwenden mechanistische Ansätze, um zu bestimmen, wie er auf zellulärer und molekularer Ebene wirkt.
Ultrastrukturelle Studien von Bakterienzellen und -gemeinschaften, die mit Honig behandelt wurden
Honig kann die Größe und Form von Bakterienzellen erheblich verändern, obwohl das Ausmaß je nach Bakterienart variiert. Mithilfe der Transmissionselektronenmikroskopie (TEM) zeigten S. aureus-Kulturen, die mit Manuka-Honig behandelt wurden, mehr Zellen mit abgeschlossenen Septen im Vergleich zu denen, die mit künstlichem Honig behandelt wurden. Dies deutet darauf hin, dass die Zellen zwar in die Teilungsphase des Zellzyklus eintraten, diese jedoch nicht abschließen konnten, obwohl diese Zellen äußerlich durch Rasterelektronenmikroskopie (REM) normal erschienen (Henriques et al., 2010). Neuere Phasenkontrastaufnahmen nach Behandlung mit einer subletalen Dosis Manuka-Honig zeigten, dass Zellen von S. aureus und Bacillus subtilis signifikant kleiner waren und eher kondensierte DNA aufwiesen als Zellen, die ohne Honig wuchsen (Lu et al., 2013). Ein direkter Vergleich dieser Studien ist schwierig, da unterschiedliche Honigmengen und Behandlungszeiten verwendet wurden, doch insgesamt deuten die Ergebnisse auf eine Entkopplung von Wachstum und Zellteilung hin, wie sie häufig als Reaktion auf ernährungsbedingte und Umweltstressfaktoren beobachtet wird (Silva-Rocha und de Lorenzo, 2010).
Die Behandlung mit Honig wurde berichtet, dass Kulturen der gramnegativen Spezies E. coli und P. aeruginosa sowohl ungewöhnlich kürzere als auch längere Zellen aufweisen (Lu et al., 2013). Interessanterweise scheint P. aeruginosa weniger anfällig für die Hemmung durch Honig zu sein als andere Spezies, dennoch wurden mit TEM und SEM tiefgreifende zelluläre Veränderungen beobachtet, darunter Furchen und Bläschen (Ausstülpungen der Zellmembranen) auf der Zelloberfläche sowie eine erhebliche Menge extrazellulärer Trümmer, die auf Zelllyse hinweisen (Henriques et al., 2011). Dies wurde in einer nachfolgenden Studie mit BacLight-Live-Dead-Fluoreszenzfärbung und Konfokalmikroskopie bestätigt, wobei jedoch auch gezeigt wurde, dass eine relativ große Anzahl lebender Zellen erhalten blieb. Diese Studien verwendeten 20 % (w/v) Honig, was höher als die MBC für ihren P. aeruginosa-Stamm war, sodass eine erhebliche Hemmung und Abtötung zu erwarten war. Dennoch fand die Rasterkraftmikroskopie (AFM) bei sub-bakteriziden Konzentrationen immer noch erhebliche Zellverzerrungen und Bläschenbildung bei Zellen, die mit MIC (12 %) und halber MIC (6 %) behandelt wurden, sowie eine erhebliche Zelllyse (Roberts et al., 2012). Diese scheinbare Degeneration der P. aeruginosa-Zelle wurde durch quantitative PCR-Analysen unterstützt, die eine zehnfache Herunterregulierung von oprF in honigbehandelten Zellen zeigten, welches ein äußeres Membranporin kodiert, das für die strukturelle Stabilität wichtig ist (Jenkins et al., 2015a).
‘Omics-Analysen bewerten die Ganzzellreaktion auf Hemmung durch Honig
Die Fähigkeit, die Gesamtausgabe von Zellen zu bewerten, hat die Untersuchung von Wechselwirkungen zwischen Arzneimitteln und Krankheitserregern revolutioniert und ist besonders wertvoll für komplexe Naturprodukte wie Honig, bei denen Effekte auf mehrere Prozesse wahrscheinlich sind. Microarray- und proteomische Studien an Bakterien, die Honig ausgesetzt waren, deuteten auf eine Induktion stressbezogener Prozesse und eine Unterdrückung der Proteinsynthese hin (Blair et al., 2009; Jenkins et al., 2011; Packer et al., 2012). Insgesamt ist dies eine recht typische Reaktion auf hemmende Mittel, doch Honig erzeugte eine einzigartige „Signatur“ differenzieller Expression, die viele Proteine mit hypothetischen oder unbekannten Funktionen einschloss, was auf einen neuartigen Wirkmechanismus hindeutet. Spezifische Gene oder Proteine, die in Omics-Analysen von S. aureus und E. coli O157/H7 herunterreguliert wurden, haben Funktionen, die mit Virulenz, Quorum Sensing und Biofilmbildung zusammenhängen (Lee et al., 2011; Jenkins et al., 2013), und bei P. aeruginosa wurde eine Herunterregulierung von Proteinen beobachtet, die an der Flagellierung beteiligt sind (Roberts et al., 2015). Diese Phänotypen sind entscheidend für Pathogene, um sich zu etablieren und invasive Infektionen zu verursachen, und zeigen, dass Honig neben der Hemmung des Wachstums auch das pathogene Potenzial infizierender Bakterien reduzieren kann.
Obwohl die bisher durchgeführten „Omics“-Analysen noch relativ begrenzt in Anzahl und Umfang sind, deuten sie auf eine komplexe zelluläre Reaktion auf Honig mit erheblicher Variation bei verschiedenen Bakterienarten hin. Fortschrittliche Systembiologie-Ansätze, die eine Kontextualisierung der Daten ermöglichen, sowie Validierungsstudien mit quantitativer PCR und Gen-Deletion-Stämmen sind nun erforderlich, um diese Komplexität zu entschlüsseln. Diese könnten neue Ansätze für Arzneimitteltherapien aufzeigen, die darauf abzielen, das Bakterienwachstum zu hemmen (Hudson et al., 2012).
Wechselwirkungen zwischen Honig und herkömmlichen Antibiotika
Neben der Verwendung als alleiniges Mittel besteht die Möglichkeit, Honig zur Ergänzung der Behandlung mit herkömmlichen Antibiotika einzusetzen. Dies kann besonders wertvoll sein, wenn es mit systemischen Mitteln kombiniert wird, die über die Blutbahn zum Wundbett gelangen, während Honig topisch angewendet wird. Kombinierte Behandlungen können auch die therapeutische Dosis antimikrobieller Mittel senken und die Entwicklung von Resistenzen verhindern; in einigen Fällen kann dies zu einer Arzneimittelsynergie führen, bei der die kombinierte Wirkung größer ist als die Summe der einzelnen Wirkungen der jeweiligen Arzneimittelpartner.
In vitro-Studien, die therapeutisch zugelassenen Manuka-Honig mit Antibiotika kombinierten, fanden eine synergistische Wirkung mit Oxacillin, Tetracyclin, Imipenem und Mupirocin gegen das Wachstum eines MRSA-Stamms (Jenkins und Cooper, 2012). Darüber hinaus stellte die Anwesenheit einer subinhibitorischen Konzentration von Honig in Kombination mit Oxacillin die Oxacillin-Empfindlichkeit des MRSA-Stamms wieder her. Die Autoren fanden eine Herunterregulierung von mecR1, das ein MRSA-spezifisches Penicillin-bindendes Protein (PBP2A) kodiert, und schlugen dies als Mechanismus der Honigsynergie vor. Eine starke synergistische Aktivität zwischen Manuka-Honig und Rifampicin gegen mehrere S. aureus-Stämme, einschließlich klinischer Isolate und MRSA-Stämme, wurde ebenfalls festgestellt, und die Anwesenheit von Honig verhinderte in vitro das Entstehen von Rifampicin-Resistenzen (Müller et al., 2013). Dies ist klinisch bedeutsam, da Rifampicin gut in Gewebe und Abszesse eindringt und häufig zur Behandlung oberflächlicher Staphylokokken-Infektionen verwendet wird, aber schnell Resistenzen induziert und daher in Kombination mit einem anderen Wirkstoff eingesetzt werden muss. Eine weitere Erkenntnis dieser Studie war, dass die Synergie nicht auf MGO zurückzuführen ist, da ein synthetischer Honig, der mit MGO versetzt wurde, keine Synergie mit Rifampicin zeigte.
Das Verständnis darüber, wie Honig die Wirkung von antimikrobiellen Mitteln mit gut charakterisierten Wirkmechanismen beeinflusst, kann auch unser Verständnis darüber vertiefen, wie Honig bakterielle Krankheitserreger beeinflusst. Liu et al. (2014) erweiterten die Analyse der Synergie um weitere Antibiotika und verschiedene S. aureus- und MRSA-Stämme. Sie vermuteten, dass eine erhöhte Empfindlichkeit gegenüber Clindamycin und Gentamicin durch die kombinierte Wirkung der durch Honig herunterregulierten Proteinsynthese und der Hemmung der Ribosomen durch die Antibiotika entstehen könnte, während die Synergie mit β-Laktam-Antibiotika auf den durch beide Partner verursachten erhöhten oxidativen Stress zurückzuführen sein könnte. Da S. aureus- und MRSA-Stämme gleichermaßen empfindlich auf die Kombination aus Oxacillin und Honig reagierten, schien es unwahrscheinlich, dass die Synergie auf eine Herunterregulierung von PBP2A zurückzuführen ist. Bei einem klinischen MRSA-Isolat wurde jedoch keine erhöhte Empfindlichkeit gegenüber Clindamycin oder Gentamicin in Anwesenheit von Honig festgestellt, was bemerkenswert ist, da dies der erste berichtete Fall einer unterschiedlichen Reaktion auf Honig bei MRSA im Vergleich zu S. aureus ist. Die Untersuchung dieses stammspezifischen Unterschieds mittels transkriptomischer oder proteomischer Analysen wäre ein interessanter Ansatz für zukünftige Forschungen (Liu et al., 2014).
Nachweise der Wirksamkeit aus Tierversuchen, Fallberichten und klinischen Studien
Unternehmen, die Manuka-Honig produzieren und vermarkten, fördern hohe ethische Standards und lehnen den Einsatz von Tiermodellen zur Untersuchung von Infektionen und Wundheilung ab. Manuka-Honig wurde jedoch zur Behandlung von Tieren mit chirurgischen oder Unfallwunden, insbesondere Pferden, mit positiven Ergebnissen eingesetzt (Dart et al., 2015; Bischofberger et al., 2016). Fallberichte über die Anwendung von Honig bei nicht heilenden Wunden und Geschwüren verzeichneten eine deutliche Verbesserung mit Infektionsauflösung, wo herkömmliche Antibiotika versagt hatten (Regulski, 2008; Smith et al., 2009). Trotz dieser Berichte und der Evidenz aus zahlreichen in vitro- und in vivo-Modellen, die zeigen, dass Honig problematische Wundpathogene abtötet, fehlen robuste klinische Daten für Manuka-Honig. Dafür gibt es verschiedene Gründe, darunter technische Schwierigkeiten bei der Durchführung einer doppelblinden, placebokontrollierten Studie mit einer charakteristischen Substanz wie Honig, ethische Überlegungen, mangelndes Interesse klinischer Praktiker und das Kosten-Nutzen-Verhältnis für Honigunternehmen, deren Fokus auf Naturprodukten und dem Verkauf über den Ladentisch liegt, wo Manuka-Honig und zugehörige Verbände bereits einen Premiumpreis erzielen. Diese Umstände könnten sich ändern, wenn die Antibiotikaresistenz aktuelle Behandlungsoptionen einschränkt und laufende Forschungen das Potenzial von Honig stärker ins Bewusstsein medizinischer Fachkräfte rücken.
Lücken und neue Chancen in der Honigforschung
In letzter Zeit wurden große Fortschritte im Verständnis von therapeutischem Honig erzielt, doch seine Anwendung in der klinischen Medizin bleibt begrenzt, selbst wenn herkömmliche Antibiotika zunehmend versagen. Die Komplexität des Honigs, die wohl seine größte Stärke bei der Abtötung vielfältiger Krankheitserreger und der Verhinderung von Resistenzen darstellt, erschwert seine Untersuchung, da viele Faktoren zusammenwirken und die Aktivität beeinflussen. Wir plädieren für weitere mechanistische Studien mit entsprechend registriertem therapeutischem Manuka-Honig, insbesondere Studien, die nicht-reduktionistische Systembiologie-Ansätze verwenden, zusammen mit detaillierten chemischen und mikrobiologischen Analysen, um zu klären, wie Honig auf molekularer, zellulärer und Populationsebene wirkt, wie dies bei verschiedenen Stämmen und Arten mikrobieller Krankheitserreger variieren kann und wie die Wirtszelle reagiert (Tabelle 2). Die aus diesen Studien gewonnenen Informationen können dann die Therapie informieren und die klinischen Daten liefern, die erforderlich sind, um Honig in die Schulmedizin zu integrieren; nicht mehr als alternative Therapie, die nur angewendet wird, wenn alle anderen Optionen versagt haben.
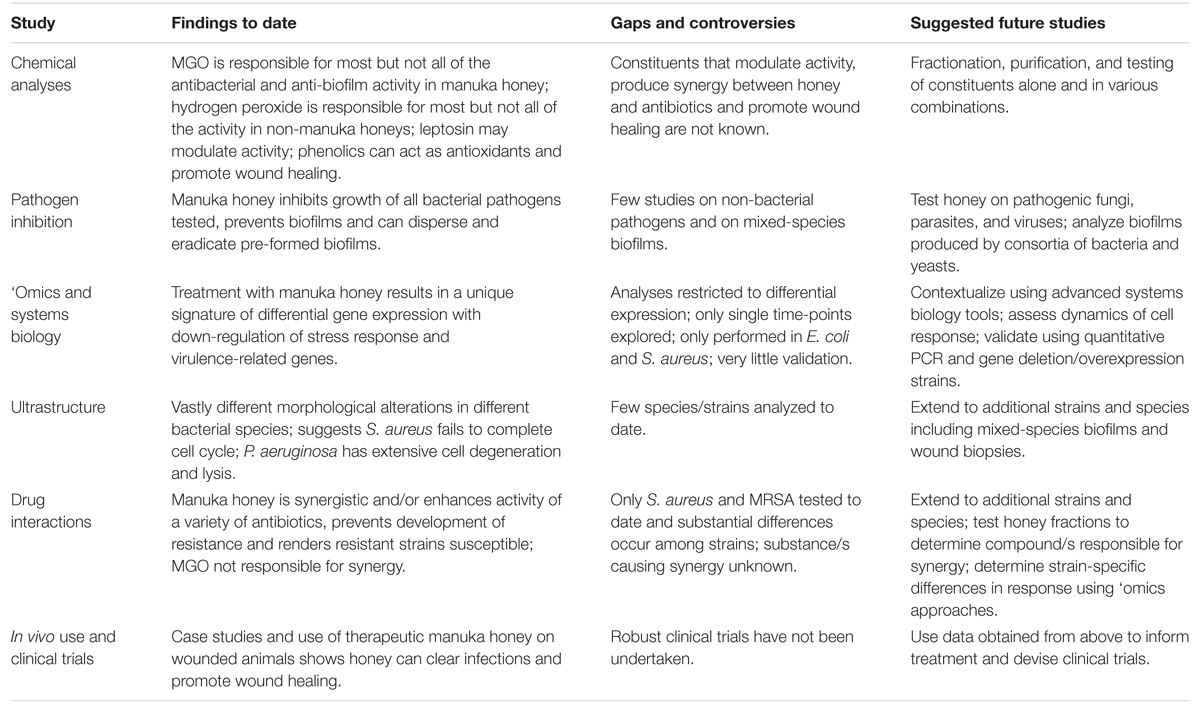
TABELLE 2. Studien zu Manuka-Honig: Erkenntnisse, Lücken und zukünftige Studien.
Autorenbeiträge
Diese Übersicht wurde von DC, SB, NNC, DB und PB verfasst und von RS und EH kritisch geprüft.
Finanzierung
NNC erhält Gehaltsunterstützung vom Rural Industries Research and Development Corporation – Honey Bee Program (Förderung PRJ-009186).
Erklärung zum Interessenkonflikt
DC, PB und EH berichten über Zuschüsse und nicht-finanzielle Unterstützung in Form von Manuka-Honig von Comvita NZ Limited und Capilano Honey Limited; RS ist bei Comvita NZ Limited angestellt, das medizinischen Manuka-Honig (Medihoney) vertreibt.
Die übrigen Autoren erklären, dass die Forschung ohne jegliche kommerzielle oder finanzielle Beziehung durchgeführt wurde, die als potenzieller Interessenkonflikt ausgelegt werden könnte.
Abkürzung
ESBL, erweitertes Spektrum β-Laktamase; MBC, minimale bakterizide Konzentration; MGO, Methylglyoxal; MIC, minimale Hemmkonzentration; MRSA, methicillinresistenter Staphylococcus aureus; MRSE, methicillinresistenter Staphylococcus epidermis; NPA, Nicht-Peroxid-Aktivität; VRE, vancomycinresistenter Enterococcus.
Literaturverzeichnis
Adams, C. J., Boult, C. H., Deadman, B. J., Farr, J. M., Grainger, M. N. C., Manley-Harris, M., et al. (2008). Isolierung mittels HPLC und Charakterisierung des bioaktiven Fraktionsanteils von neuseeländischem Manuka (Leptospermum scoparium) Honig. Carbohydr. Res. 343, 651–659. doi: 10.1016/j.carres.2007.12.011
Adams, C. J., Manley-Harris, M., und Molan, P. C. (2009). Die Herkunft von Methylglyoxal in neuseeländischem Manuka (Leptospermum scoparium) Honig. Carbohydr. Res. 344, 1050–1053. doi: 10.1016/j.carres.2009.03.020
Al Somal, N., Coley, K. E., Molan, P. C., und Hancock, B. M. (1994). Empfindlichkeit von Helicobacter pylori gegen die antibakterielle Aktivität von Manuka-Honig. J. R. Soc. Med. 87, 9–12.
Allen, K., Molan, P., und Reid, G. (1991). Eine Untersuchung der antibakteriellen Aktivität einiger neuseeländischer Honige. J. Pharm. Pharmacol.43, 817–822. doi: 10.1111/j.2042-7158.1991.tb03186.x
Allen, K. L., und Molan, P. C. (1997). Die Empfindlichkeit von Mastitis-verursachenden Bakterien gegenüber der antibakteriellen Wirkung von Honig. N. Z. J. Agric. Res. 40, 537–540. doi: 10.1080/00288233.1997.9513276
Anthimidou, E., und Mossialos, D. (2012). Antibakterielle Aktivität griechischer und zyprischer Honige gegen Staphylococcus aureus und Pseudomonas aeruginosa im Vergleich zu Manuka-Honig. J. Med. Food 16, 42–47. doi: 10.1089/jmf.2012.0042
Balan, P., Mal, G., Das, S., und Singh, H. (2016). Synergistische und additive antimikrobielle Wirkungen von Curcumin, Manuka-Honig und Molkenproteinen. J. Food Biochem. doi: 10.1111/jfbc.12249
Bassam, Z., Zohra, B. I., und Saada, A.-A. (1997). Die Wirkung von Honig auf Leishmania-Parasiten: eine in vitro-Studie. Trop. Doctor27, 36–38.
Biglari, B., Moghaddam, A., Santos, K., Blaser, G., Büchler, A., Jansen, G., et al. (2013). Multizentrische prospektive Beobachtungsstudie zur professionellen Wundversorgung mit Honig (Medihoney). Int. Wound J. 10, 252–259. doi: 10.1111/j.1742-481X.2012.00970.x
Bischofberger, A., Dart, C., Horadagoda, N., Perkins, N., Jeffcott, L., Little, C., et al. (2016). Wirkung von Manuka-Honig-Gel auf die Konzentrationen von Transforming Growth Factor β1 und β3, Bakterienzahlen und Histomorphologie kontaminierter Hautwunden voller Dicke an den distalen Gliedmaßen von Pferden. Aust. Vet. J. 94, 27–34. doi: 10.1111/avj.12405
Blair, S., Cokcetin, N., Harry, E., und Carter, D. (2009). Die ungewöhnliche antibakterielle Aktivität von medizinischem Leptospermum-Honig: antibakterielles Spektrum, Resistenz und Transkriptomanalyse. Eur. J. Clin. Microbiol. Infect. Dis. 28, 1199–1208. doi: 10.1007/s10096-009-0763-z
Blair, S. E., und Carter, D. A. (2005). Das Potenzial von Honig bei der Behandlung von Wunden und Infektionen. J. Austral. Infect. Control 10, 24–31.
Brady, N., Molan, P., und Harfoot, C. (1996). Die Empfindlichkeit von Dermatophyten gegenüber der antimikrobiellen Aktivität von Manuka-Honig und anderem Honig. Pharm. Pharmacol. Commun. 2, 471–473.
Carnwath, R., Graham, E. M., Reynolds, K., und Pollock, P. J. (2014). Die antimikrobielle Aktivität von Honig gegen häufige bakterielle Isolate aus Pferdewunden. Vet. J. 199, 110–114. doi: 10.1016/j.tvjl.2013.07.003
Chen, C., Campbell, L., Blair, S. E., und Carter, D. A. (2012). Die Auswirkung von Hitzebehandlung auf die antimikrobiellen Eigenschaften von Honig. Front. Microbiol. 3:265. doi: 10.3389/fmicb.2012.00265
Cooper, R., Jenkins, L., Henriques, A., Duggan, R., und Burton, N. (2010). Keine bakterielle Resistenz gegen medizinischen Manuka-Honig. Eur. J. Clin. Microbiol. Infect. Dis. 29, 1237–1241. doi: 10.1007/s10096-010-0992-1
Cooper, R. A., Halas, E., und Molan, P. C. (2002a). Die Wirksamkeit von Honig bei der Hemmung von Stämmen von Pseudomonas aeruginosa aus infizierten Verbrennungen. J. Burn Care Rehabil. 23, 366–370. doi: 10.1097/00004630-200211000-00002
Cooper, R. A., und Molan, P. C. (1999). Die Verwendung von Honig als Antiseptikum bei der Behandlung von Pseudomonas-Infektionen. J. Wound Care 8, 161–164. doi: 10.12968/jowc.1999.8.4.25867
Cooper, R. A., Molan, P. C., und Harding, K. G. (1999). Antibakterielle Aktivität von Honig gegen Stämme von Staphylococcus aureus aus infizierten Wunden. J. R. Soc. Med. 92, 283–285.
Cooper, R. A., Molan, P. C., und Harding, K. G. (2002b). Die Empfindlichkeit klinisch relevanter grampositiver Kokken aus Wunden gegenüber Honig. J. Appl. Microbiol. 93, 857–863. doi: 10.1046/j.1365-2672.2002.01761.x
Cooper, R. A., Wigley, P., und Burton, N. F. (2000). Empfindlichkeit multiresistenter Stämme von Burkholderia cepacia gegen Honig. Lett. Appl. Microbiol. 31, 20–24. doi: 10.1046/j.1472-765x.2000.00756.x
Dart, A., Bischofberger, A., Dart, C., und Jeffcott, L. (2015). Eine Übersicht über die Forschung zur Wundheilung zweiter Absicht bei Pferden mit Manuka-Honig: aktuelle Empfehlungen und zukünftige Anwendungen. Equine Vet. Educ. 27, 658–664. doi: 10.1111/eve.12379
Estevinho, L., Pereira, A. P., Moreira, L., Dias, L. G., und Pereira, E. (2008). Antioxidative und antimikrobielle Wirkungen von phenolischen Verbindungen aus Honig aus Nordostportugal. Food Chem. Toxicol. 46, 3774–3779. doi: 10.1016/j.fct.2008.09.062
French, V. M., Cooper, R. A., und Molan, P. C. (2005). Die antibakterielle Aktivität von Honig gegen koagulase-negative Staphylokokken. J. Antimikrobielle Chemother. 56, 228–231. doi: 10.1093/jac/dki193
George, N. M., und Cutting, K. F. (2007). Antibakterieller Honig (Medihoney): in-vitro-Aktivität gegen klinische Isolate von MRSA, VRE und anderen multiresistenten gramnegativen Organismen einschließlich Pseudomonas aeruginosa. Wunden 19:231.
Halstead, F. D., Webber, M. A., Rauf, M., Burt, R., Dryden, M., und Oppenheim, B. A. (2016). In vitro-Aktivität eines entwickelten Honigs, medizinischer Honige und antimikrobieller Wundauflagen gegen biofilmbildende klinische Bakterienisolaten. J. Wundversorgung 25, 93–102. doi: 10.12968/jowc.2016.25.2.93
Hammond, E. N., und Donkor, E. S. (2013). Antibakterielle Wirkung von Manuka-Honig auf Clostridium difficile. BMC Res. 6:188. doi: 10.1186/1756-0500-6-188
Henriques, A. F., Jenkins, R. E., Burton, N. F., und Cooper, R. A. (2010). Die intrazellulären Effekte von Manuka-Honig auf Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 29, 45–50. doi: 10.1007/s10096-009-0817-2
Henriques, A. F., Jenkins, R. E., Burton, N. F., und Cooper, R. A. (2011). Die Wirkung von Manuka-Honig auf die Struktur von Pseudomonas aeruginosa. Eur. J. Clin. Microbiol. Infect. Dis. 30, 167–171. doi: 10.1007/s10096-010-1065-1
Hudson, N. J., Dalrymple, B. P., und Reverter, A. (2012). Über die differentielle Expression hinaus: die Suche nach kausalen Mutationen und Effektormolekülen. BMC Genomics 13:356. doi: 10.1186/1471-2164-13-356
Irish, J., Blair, S., und Carter, D. (2011). Die antibakterielle Aktivität von Honig, der aus australischer Flora gewonnen wurde. PLoS ONE 6:e18229. doi: 10.1371/journal.pone.0018229
Irish, J., Carter, D. A., Shokohi, T., und Blair, S. E. (2006). Honig hat eine antifungale Wirkung gegen Candida-Arten. Med. Mycol. 44, 289–291. doi: 10.1080/13693780600931986
Jenkins, R., Burton, N., und Cooper, R. (2011). Wirkung von Manuka-Honig auf die Expression des universellen Stressproteins A bei methicillinresistentem Staphylococcus aureus. Int. J. Antimicrob. Agents 37, 373–376. doi: 10.1016/j.ijantimicag.2010.11.036
Jenkins, R., Burton, N., und Cooper, R. (2013). Proteomische und genomische Analyse von methicillinresistentem Staphylococcus aureus (MRSA), das in vitro Manuka-Honig ausgesetzt wurde, zeigte eine Herunterregulierung von Virulenzmarkern. J. Antimicrobial Chemother. 69, 603–615. doi: 10.1093/jac/dkt430
Jenkins, R., und Cooper, R. (2012). Verbesserung der Antibiotika-Wirkung gegen Wundpathogene mit Manuka-Honig in vitro. PLoS ONE 7:e45600. doi: 10.1371/journal.pone.0045600
Jenkins, R., Roberts, A., und Brown, H. L. (2015a). Über die antibakteriellen Effekte von Manuka-Honig: mechanistische Einblicke. Res. Rep. Biol. 6, 215–224. doi: 10.2147/RRB.S75754
Jenkins, R., Wootton, M., Howe, R., und Cooper, R. (2015b). Eine Demonstration der Empfindlichkeit klinischer Isolate von Mukoviszidose-Patienten gegenüber Manuka-Honig. Arch. Microbiol. 197, 597–601. doi: 10.1007/s00203-015-1091-6
Kalapos, M. P. (2008). Das Tandem von freien Radikalen und Methylglyoxal. Chem. Biol. Interact. 171, 251–271. doi: 10.1016/j.cbi.2007.11.009
Kato, Y., Umeda, N., Maeda, A., Matsumoto, D., Kitamoto, N., und Kikuzaki, H. (2012). Identifizierung eines neuartigen Glykosids, Leptosin, als chemischer Marker für Manuka-Honig. J. Agric. Food Chem. 60, 3418–3423. doi: 10.1021/jf300068w
Kilty, S. J., Duval, M., Chan, F. T., Ferris, W., und Slinger, R. (2011). Methylglyoxal: (aktiver Wirkstoff des Manuka-Honigs) in vitro-Aktivität gegen bakterielle Biofilme. Int. Forum Allergy Rhinol. 1, 348–350. doi: 10.1002/alr.20073
Kronda, J. M., Cooper, R. A., und Maddocks, S. E. (2013). Manuka-Honig hemmt die Siderophorproduktion bei Pseudomonas aeruginosa. J. Appl. Microbiol. 115, 86–90. doi: 10.1111/jam.12222
Kwakman, P. H., te Velde, A. A., de Boer, L., Vandenbroucke-Grauls, C. M., und Zaat, S. A. (2011). Zwei wichtige medizinische Honige haben unterschiedliche Mechanismen der bakteriziden Wirkung. PLoS ONE 6:e17709. doi: 10.1371/journal.pone.0017709
Lee, J.-H., Park, J.-H., Kim, J.-A., Neupane, G. P., Cho, M. H., Lee, C.-S., et al. (2011). Niedrige Konzentrationen von Honig reduzieren die Biofilmbildung, Quorum Sensing und Virulenz bei Escherichia coli O157: H7. Biofouling 27, 1095–1104. doi: 10.1080/08927014.2011.633704
Lin, S. M., Molan, P. C., und Cursons, R. T. (2011). Die kontrollierte in vitro Empfindlichkeit gastrointestinaler Krankheitserreger gegenüber der antibakteriellen Wirkung von Manuka-Honig. Eur. J. Clin. Microbiol. Infect. Dis. 30, 569–574. doi: 10.1007/s10096-010-1121-x
Liu, M., Lu, J., Müller, P., Turnbull, L., Burke, C. M., Schlothauer, R. C., et al. (2014). Antibiotika-spezifische Unterschiede in der Reaktion von Staphylococcus aureus auf die Behandlung mit antimikrobiellen Mitteln in Kombination mit Manuka-Honig. Front. Microbiol. 5:779. doi: 10.3389/fmicb.2014.00779
Lu, J., Carter, D. A., Turnbull, L., Rosendale, D., Hedderley, D., Stephens, J., et al. (2013). Die Wirkung von neuseeländischem Kanuka-, Manuka- und Kleehonig auf das bakterielle Wachstum und die Zellmorphologie variiert je nach Art. PLoS ONE8:e55898. doi: 10.1371/journal.pone.0055898
Lu, J., Turnbull, L., Burke, C. M., Liu, M., Carter, D. A., Schlothauer, R. C., et al. (2014). Manuka-Honige können Biofilme von Staphylococcus aureus-Stämmen mit unterschiedlichen Biofilm-Bildungsfähigkeiten beseitigen. PeerJ 2:e326. doi: 10.7717/peerj.326
Maddocks, S. E., Jenkins, R. E., Rowlands, R. S., Purdy, K. J., und Cooper, R. A. (2013). Manuka-Honig hemmt die Adhäsion und Invasion medizinisch wichtiger Wundbakterien in vitro. Fut. Microbiol. 8, 1523–1536. doi: 10.2217/fmb.13.126
Maddocks, S. E., Lopez, M. S., Rowlands, R. S., und Cooper, R. A. (2012). Manuka-Honig hemmt die Entwicklung von Streptococcus pyogenes-Biofilmen und verursacht eine reduzierte Expression von zwei Fibronectin-bindenden Proteinen. Microbiology 158, 781–790. doi: 10.1099/mic.0.053959-0
Majtan, J. (2014). Honig: ein Immunmodulator bei der Wundheilung. Wundheilung Regenerat. 22, 187–192. doi: 10.1111/wrr.12117
Majtan, J., Bohova, J., Horniackova, M., Klaudiny, J., und Majtan, V. (2014a). Anti-Biofilm-Wirkungen von Honig gegen Wundpathogene Proteus mirabilis und Enterobacter cloacae. Phytother. Res. 28, 69–75. doi: 10.1002/ptr.4957
Majtan, J., Bohova, J., Prochazka, E., und Klaudiny, J. (2014b). Methylglyoxal kann die Wasserstoffperoxid-Akkumulation im Manuka-Honig durch Hemmung der Glukoseoxidase beeinflussen. J. Med. Food 17, 290–293. doi: 10.1089/jmf.2012.0201
Majtan, J., Klaudiny, J., Bohova, J., Kohutova, L., Dzurova, M., Sediva, M., et al. (2012). Methylglyoxal-induzierte Modifikationen bedeutender proteinartiger Komponenten der Honigbiene im Manuka-Honig: mögliche therapeutische Implikationen. Fitoterapia 83, 671–677. doi: 10.1016/j.fitote.2012.02.002
Majtan, J., Majtanova, L., Bohova, J., und Majtan, V. (2011). Honigtauhonig als potentes antibakterielles Mittel zur Ausrottung multiresistenter Stenotrophomonas maltophilia-Isolate von Krebspatienten. Phytother. Res. 25, 584–587. doi: 10.1002/ptr.3304
Malone, L. A., Gatehouse, H. S., und Tregidga, E. L. (2001). Auswirkungen von Zeit, Temperatur und Honig auf Nosema apis (Microsporidia: Nosematidae), einen Parasiten der Honigbiene, Apis mellifera (Hymenoptera: Apidae). J. Invertebrate Pathol. 77, 258–268. doi: 10.1006/jipa.2001.5028
Mavric, E., Wittmann, S., Barth, G., und Henle, T. (2008). Identifikation und Quantifizierung von Methylglyoxal als dominierendem antibakteriellem Bestandteil von Manuka (Leptospermum scoparium) Honigen aus Neuseeland. Mol. Nutrit. Food Res. 52, 483–489. doi: 10.1002/mnfr.200700282
Molan, P. M. (2008). Eine Erklärung, warum der MGO-Gehalt in Manuka-Honig die antibakterielle Aktivität nicht zeigt. New Zealand Beekeeper 16, 11–13.
Mullai, V., und Menon, T. (2007). Bakterizide Aktivität verschiedener Honigsorten gegen klinische und Umweltisolaten von Pseudomonas aeruginosa. J. Alternat. Complement. Med. 13, 439–442. doi: 10.1089/acm.2007.6366
Müller, P., Alber, D. G., Turnbull, L., Schlothauer, R. C., Carter, D. A., Whitchurch, C. B., et al. (2013). Synergismus zwischen Medihoney und Rifampicin gegen methicillinresistente Staphylococcus aureus (MRSA). PLoS ONE 8:e57679. doi: 10.1371/journal.pone.0057679
Mundo, M. A., Padilla-Zakour, O. I., und Worobo, R. W. (2004). Wachstumshemmung von lebensmittelbedingten Krankheitserregern und lebensmittelverderbenden Organismen durch ausgewählte Rohhonige. Int. J. Food Microbiol. 97, 1–8. doi: 10.1016/j.ijfoodmicro.2004.03.025
Nilforoushzadeh, M. A., Jaffary, F., Moradi, S., Derakhshan, R., und Haftbaradaran, E. (2007). Wirkung der topischen Honiganwendung zusammen mit intraläsionaler Glucantime-Injektion bei der Behandlung der kutanen Leishmaniose. BMC Complement Altern. Med. 7:1. doi: 10.1186/1472-6882-7-1
Norton, A. M., McKenzie, L. N., Brooks, P. R., und Pappalardo, L. J. (2015). Quantifizierung von Dihydroxyaceton im australischen Leptospermum-Nektar mittels Hochleistungsflüssigkeitschromatographie. J. Agric. Food Chem. 63, 6513–6517. doi: 10.1021/acs.jafc.5b01930
Okhiria, O., Henriques, A., Burton, N., Peters, A., und Cooper, R. (2009). Honig moduliert Biofilme von Pseudomonas aeruginosa zeit- und dosisabhängig. J. ApiProduct. ApiMedical Sci. 1, 6–10. doi: 10.3896/IBRA.4.01.1.03
Osato, M. S., Reddy, S. G., und Graham, D. Y. (1999). Osmotische Wirkung von Honig auf Wachstum und Lebensfähigkeit von Helicobacter pylori. Dig. Dis. Sci. 44, 462–464. doi: 10.1023/A:1026676517213
Packer, J. M., Irish, J., Herbert, B. R., Hill, C., Padula, M., Blair, S. E., et al. (2012). Spezifische nicht-peroxidische antibakterielle Wirkung von Manuka-Honig auf das Staphylococcus aureus-Proteom. Int. J. Antimicrob. Agents 40, 43–50. doi: 10.1016/j.ijantimicag.2012.03.012
Regulski, M. (2008). Ein neuartiger Wundverband für chronische Beingeschwüre. Podiatry Manag. 27, 235–246.
Roberts, A. E., Maddocks, S. E., und Cooper, R. A. (2012). Manuka-Honig ist bakterizid gegen Pseudomonas aeruginosa und führt zu einer differenzierten Expression von oprF und algD. Microbiology 158, 3005–3013. doi: 10.1099/mic.0.062794-0
Roberts, A. E. L., Maddocks, S. E., und Cooper, R. A. (2015). Manuka-Honig reduziert die Beweglichkeit von Pseudomonas aeruginosa durch Unterdrückung von Flagellen-assoziierten Genen. J. Antimicrob. Chemother. 70, 716–725. doi: 10.1093/jac/dku448
Sajid, M., und Azim, M. K. (2012). Charakterisierung der nematiziden Aktivität von natürlichem Honig. J. Agric. Food Chem. 60, 7428–7434. doi: 10.1021/jf301653n
Shahzad, A., und Cohrs, R. J. (2012). In vitro antivirale Aktivität von Honig gegen Varizella-Zoster-Virus (VZV): eine translationalmedizinische Studie für ein potenzielles Mittel gegen Gürtelrose. Transl. Biomed. 3:2.
Sherlock, O., Dolan, A., Athman, R., Power, A., Gethin, G., Cowman, S., et al. (2010). Vergleich der antimikrobiellen Aktivität von Ulmo-Honig aus Chile und Manuka-Honig gegen methicillinresistente Staphylococcus aureus, Escherichia coli und Pseudomonas aeruginosa. BMC Complement Altern. Med. 10:47. doi: 10.1186/1472-6882-10-47
Silva-Rocha, R., und de Lorenzo, V. (2010). Rauschen und Robustheit in prokaryotischen Regulationsnetzwerken. Annu. Rev. Microbiol. 64, 257–275. doi: 10.1146/annurev.micro.091208.073229
Smith, T., Hanft, J. R., und Legel, K. (2009). Topischer Leptospermum-Honig bei therapieresistenten venösen Beingeschwüren: eine vorläufige Fallserie. Adv. Skin Wound Care 22, 68–71. doi: 10.1097/01.ASW.0000345283.05532.9a
Soffer, A. (1976). Chihuahuas und Laetrile, Chelat-Therapie und Honig aus Boulder, Colorado [editorial]. Arch. Intern. Med.136, 865–866. doi: 10.1001/archinte.136.8.865
Sojka, M., Valachova, I., Bucekova, M., und Majtan, J. (2016). Antibiofilm-Wirksamkeit von Honig und bienenabgeleitetem Defensin-1 auf multizellulären Wundbiofilm. J. Med. Microbiol. doi: 10.1099/jmm.0.000227 [Epub ahead of print].
Stephens, J. M., Schlothauer, R. C., Morris, B. D., Yang, D., Fearnley, L., Greenwood, D. R., et al. (2010). Phenolische Verbindungen und Methylglyoxal in einigen neuseeländischen Manuka- und Kanuka-Honigen. Food Chem. 120, 78–86. doi: 10.1016/j.foodchem.2009.09.074
Tan, H. T., Rahman, R. A., Gan, S. H., Halim, A. S., Hassan, S. A., Sulaiman, S. A., et al. (2009). Die antibakteriellen Eigenschaften des malaysischen Tualang-Honigs gegen Wund- und enterische Mikroorganismen im Vergleich zu Manuka-Honig. BMC Complement Altern. Med. 9:34. doi: 10.1186/1472-6882-9-34
Watanabe, K., Rahmasari, R., Matsunaga, A., Haruyama, T., und Kobayashi, N. (2014). Anti-Influenza-Viraleffekte von Honig in vitro: starke hohe Aktivität von Manuka-Honig. Arch. Med. Res. 45, 359–365. doi: 10.1016/j.arcmed.2014.05.006
Wilkinson, J. M., und Cavanagh, H. M. A. (2005). Antibakterielle Aktivität von 13 Honigen gegen Escherichia coli und Pseudomonas aeruginosa. J. Med. Food 8, 100–103. doi: 10.1089/jmf.2005.8.100
Williams, S., King, J., Revell, M., Manley-Harris, M., Balks, M., Janusch, F., et al. (2014). Regionale, jährliche und individuelle Schwankungen im Dihydroxyaceton-Gehalt des Nektars von Manuka (Leptospermum scoparium) in Neuseeland. J. Agric. Food Chem. 62, 10332–10340. doi: 10.1021/jf5045958
Willix, D. J., Molan, P. C., und Harfoot, C. G. (1992). Ein Vergleich der Empfindlichkeit von wundeninfizierenden Bakterienarten gegenüber der antibakteriellen Aktivität von Manuka-Honig und anderem Honig. J. Appl. Bacteriol. 73, 388–394. doi: 10.1111/j.1365-2672.1992.tb04993.x
Windsor, S., Pappalardo, M., Brooks, P., Williams, S., und Manley-Harris, M. (2012). Eine praktische neue Analyse von Dihydroxyaceton und Methylglyoxal angewandt auf australische Leptospermum-Honige. J. Pharmacogn. Phytother. 4, 6–11.
Therapeutischer Manuka-Honig: Nicht mehr so alternativ - https://www.frontiersin.org/articles/10.3389/fmicb.2016.00569/full
Hinterlassen Sie einen Kommentar