عسل Manuka العلاجي: لم يعد بديلاً غريبًا

يشهد البحث في العسل الطبي نهضة كبيرة. من كونه علاجًا شعبيًا كان يُرفض إلى حد كبير من قبل الطب التقليدي باعتباره "بديلًا"، نرى الآن اهتمامًا متزايدًا من العلماء والممارسين السريريين والجمهور العام في الاستخدامات العلاجية للعسل. هناك عدة عوامل تحفز هذا الاهتمام: أولاً، ارتفاع مقاومة المضادات الحيوية لدى العديد من مسببات الأمراض البكتيرية دفع إلى تطوير واستخدام مضادات بكتيرية جديدة؛ ثانيًا، أظهرت عدد متزايد من الدراسات والتقارير الموثوقة أن بعض أنواع العسل فعالة جدًا في علاج الجروح؛ ثالثًا، العسل العلاجي يحظى بسعر مرتفع، وصناعة العسل تروج بنشاط للدراسات التي تسمح لها بالاستفادة من ذلك؛ وأخيرًا، الطبيعة المعقدة وغير المتوقعة للعسل تشكل تحديًا جذابًا للعلماء في المختبرات. في هذه الورقة نستعرض أبحاث عسل المانوكا، بدءًا من الدراسات الرصدية حول تأثيراته المضادة للميكروبات وصولًا إلى الأعمال التجريبية والآلية الحالية التي تهدف إلى إدخال العسل في الطب التقليدي. نوضح الفجوات الحالية والجدالات المتبقية في معرفتنا بكيفية عمل العسل، ونقترح دراسات جديدة قد تجعل العسل بديلًا لم يعد "بديلًا" فقط.
مقدمة
لقد استُخدم العسل كدواء طوال تاريخ البشرية. وكان من أكثر الاستخدامات العلاجية شيوعًا واستمرارية للعسل هو كضماد للجروح، ويرجع ذلك على الأرجح إلى خصائصه المضادة للميكروبات. مع ظهور المضادات الحيوية الفعالة في الستينيات، تم تجاهل العسل واعتُبر "مادة عديمة القيمة ولكنها غير ضارة" (Soffer, 1976). ومع ذلك، فإن الأزمة الحالية والمتزايدة لمقاومة المضادات الحيوية أعادت الاهتمام باستخدام العسل، سواء كعامل فعال بحد ذاته أو كدليل علاجي لتطوير طرق علاج جديدة. عادةً ما يُستخرج العسل من رحيق الأزهار وينتجه النحل، وغالبًا ما يكون نحل العسل الأوروبي Apis mellifera، وهو مزيج معقد من السكريات والأحماض الأمينية والفينولات ومواد أخرى. تختلف أنواع العسل المستخرجة من نباتات مزهرة مختلفة بشكل كبير في قدرتها على قتل البكتيريا، وقد أدى ذلك إلى تعقيد الأدبيات المتعلقة بالعسل وجعل من الصعب أحيانًا تكرار النتائج عبر دراسات مختلفة (Allen et al., 1991; Irish et al., 2011). ركزت الغالبية العظمى من الدراسات الحديثة التي تحقق في آلية عمل العسل على عسل Manuka النشط الموحد والمُعرف جيدًا، والذي يُنتج من بعض أنواع Leptospermum الأصلية في نيوزيلندا وأستراليا، وقد تم تسجيله كمنتج للعناية بالجروح لدى الهيئات التنظيمية الطبية المختصة. لذلك، ما لم يُذكر خلاف ذلك، سيركز هذا الاستعراض على عسل Manuka.
التحليلات الكيميائية لعسل Manuka النشط
كان البروفيسور بيتر مولان من جامعة وايكاتو في نيوزيلندا أول من أبلغ عن النشاط غير المعتاد لعسل المانوكا وبدأ في اختبار تأثيره ضد مجموعة واسعة من أنواع البكتيريا المختلفة في منتصف الثمانينيات. ومع ذلك، بينما كان من الواضح أن حتى التركيزات المنخفضة من عسل المانوكا تقتل مسببات الأمراض البكتيرية، ظل المكون النشط المحدد المسؤول عن ذلك غامضًا لسنوات عديدة. يجعل السكر العالي ودرجة الحموضة المنخفضة العسل مثبطًا لنمو الميكروبات، لكن النشاط يبقى حتى عند تخفيف هذه العوامل إلى مستويات ضئيلة. تنتج العديد من أنواع العسل أيضًا بيروكسيد الهيدروجين عندما يتفاعل إنزيم جلوكوز أوكسيداز، المشتق من نحلة العسل، مع الجلوكوز والماء. ومع ذلك، في عسل المانوكا يكون إنتاج بيروكسيد الهيدروجين منخفضًا نسبيًا ويمكن تحييده بواسطة إنزيم الكاتالاز، ومع ذلك يبقى النشاط. تم الكشف أخيرًا عن سبب هذا النشاط المتبقي، الذي أُطلق عليه اسم "النشاط غير البيروكسيدي" أو NPA، في عام 2008، عندما حدد مختبران بشكل مستقل الميثيل جلايوكسال (MGO) في عسل المانوكا (Adams et al., 2008; Mavric et al., 2008). ينتج MGO عن الجفاف التلقائي لمركبه السابق ديهيدروكسي أسيتون (DHA)، وهو مركب نباتي طبيعي يوجد في رحيق أزهار Leptospermum scoparium, Leptospermum polygalifolium وبعض أنواع Leptospermum ذات الصلة التي تنمو في نيوزيلندا وأستراليا (Adams et al., 2009; Williams et al., 2014; Norton et al., 2015). يمكن لـ MGO أن يتفاعل بشكل غير محدد نسبيًا مع الجزيئات الكبيرة مثل DNA وRNA والبروتينات (Adams et al., 2008; Mavric et al., 2008; Majtan et al., 2014b)، ومن الناحية النظرية قد يكون سامًا للخلايا الثديية (Kalapos, 2008). ومع ذلك، لا توجد أدلة على تلف خلايا المضيف عند استهلاك عسل المانوكا عن طريق الفم أو استخدامه كضماد للجروح؛ بل يبدو أن العسل يحفز الشفاء ويقلل من التندب عند تطبيقه على الجروح (Biglari et al., 2013; Majtan, 2014; Dart et al., 2015). ولا يُعرف كيف يمارس هذا السمية الانتقائية الظاهرة تجاه الخلايا البكتيرية.
عادةً ما تنتج المستويات العالية من MGO أو بيروكسيد الهيدروجين العسل الأكثر نشاطًا، ومع ذلك، فإن الارتباط ليس دائمًا مثاليًا مما يشير إلى أن مكونات أخرى في العسل قد تؤثر على النشاط (Molan, 2008; Kwakman et al., 2011; Chen et al., 2012; Lu et al., 2013). الديفنسين-1، وهو ببتيد مضاد للميكروبات مشتق من النحل، مسؤول عن النشاط في عسل Revamil، وهو عسل نشط يُنتج من مصدر غير معلن، لكن يبدو أنه معدل هيكليًا وغير نشط في عسل Manuka (Kwakman et al., 2011; Majtan et al., 2012). يرتبط مستوى الليبتوسين، وهو جليكوسيد يوجد حصريًا في عسل Leptospermum، بالقوة وقد يؤثر على النشاط المضاد للميكروبات في عسل Manuka (Kato et al., 2012). وبالمثل، يمكن أن تتواجد مركبات فينولية مختلفة ذات نشاط مضاد للميكروبات محتمل، خاصة في العسل ذي اللون الداكن، وعلى الرغم من أن هذه المركبات توجد بمستويات من غير المرجح أن تكون مثبطة بمفردها، إلا أنها قد تتآزر مع بعضها البعض أو مع مكونات أخرى في العسل لإنتاج أو تعديل النشاط (Estevinho et al., 2008; Stephens et al., 2010). يمكن أن تعمل الفينولات أيضًا كمضادات للأكسدة وقد تكون مسؤولة عن الخصائص المضادة للالتهابات والمساعدة في شفاء الجروح في العسل (Stephens et al., 2010). يجب ملاحظة أن ليس كل أنواع Leptospermum تنتج عسلًا نشطًا، وحتى داخل عسل L. scoparium وL. polygalifolium يمكن أن تتراوح مستويات MGO من حوالي 100 إلى أكثر من 1200 جزء في المليون (Windsor et al., 2012). أظهرت دراسة لنشاط العسل الأسترالي أن العسل المستخرج من نباتات Leptospermum التي تنمو حول حدود نيو ساوث ويلز وكوينزلاند كان نشطًا بشكل خاص، لكن ما إذا كان ذلك بسبب النبات أو التربة أو المناخ أو عوامل أخرى غير معروف (Irish et al., 2011).
تثبيط الممرضات بواسطة العسل
تم اختبار العسل مختبريًا على مجموعة متنوعة من الممرضات، خاصة تلك التي يمكن أن تستعمر الجلد والجروح والأغشية المخاطية، حيث يكون العلاج الموضعي بالعسل ممكنًا. حتى الآن، وجدت الاختبارات مختبريًا أن عسل المانوكا يمكنه تثبيط جميع الممرضات البكتيرية المسببة للمشاكل التي تم اختبارها بفعالية (ملخص في الجدول 1). ومن المثير للاهتمام بشكل خاص أن العزلات السريرية التي تحمل أنماط مقاومة متعددة للأدوية (MDR) لا تظهر انخفاضًا في حساسيتها للعسل، مما يشير إلى طيف واسع من الفعالية يختلف عن أي مضاد ميكروبي معروف (Willix et al., 1992; Blair and Carter, 2005; George and Cutting, 2007; Tan et al., 2009). بالإضافة إلى ذلك، لم تنجح المحاولات في المختبر لتوليد سلالات مقاومة للعسل، ولم ترد تقارير عن عزلات سريرية اكتسبت مقاومة للعسل (Blair et al., 2009; Cooper et al., 2010).
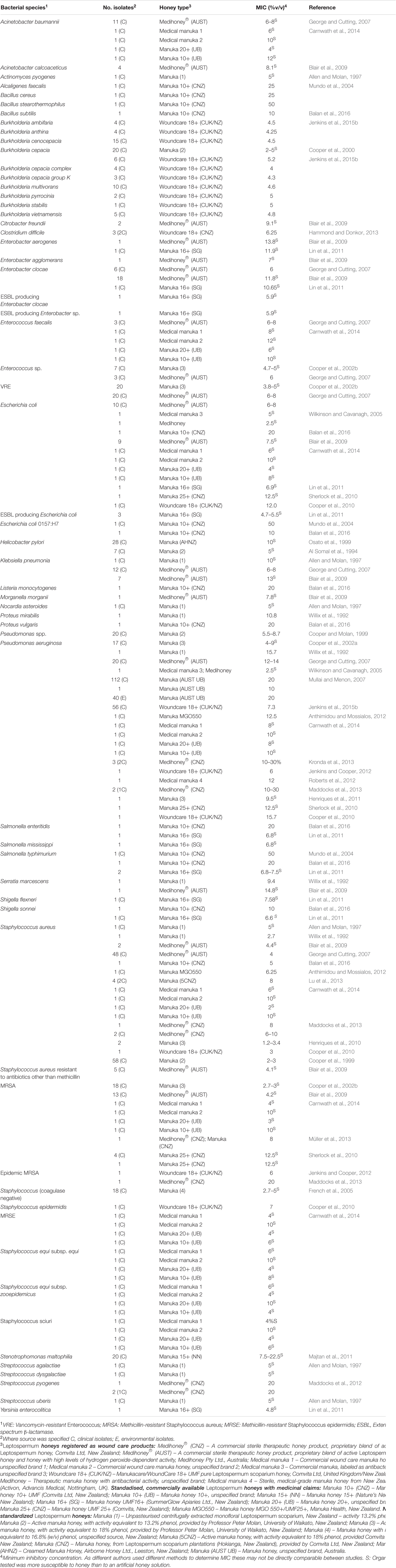
الجدول 1. الأنواع البكتيرية التي ثبت حساسيتها لعسل المانوكا العلاجي.
بالإضافة إلى تثبيط الخلايا العوالقية، يمكن للعسل أن يفرق ويقتل البكتيريا التي تعيش في الأغشية الحيوية. الأغشية الحيوية هي مجتمعات من الخلايا تكون عادةً محاطة بمصفوفة خارج خلوية تنتجها ذاتياً وتلتصق بالأسطح، بما في ذلك الجروح، الأسنان، الأسطح المخاطية، والأجهزة المزروعة. الميكروبات المقيمة في الأغشية الحيوية محمية من العوامل المضادة للميكروبات ويمكن أن تسبب التهابات مستمرة وغير قابلة للشفاء. يعطل عسل Manuka تجمعات الخلايا (Maddocks et al., 2012; Roberts et al., 2012) ويمنع تكوين الأغشية الحيوية لمجموعة واسعة من مسببات الأمراض المزعجة، بما في ذلك أنواع Streptococcus وStaphylococcus، وPseudomonas aeruginosa, Escherichia coli, Proteus mirabilis, Enterobacter cloacae, Acinetobacter baumannii، وKlebsiella pneumonia (Maddocks et al., 2012, 2013; Lu et al., 2014; Majtan et al., 2014a; Halstead et al., 2016). ومن المهم أن العسل يمكنه أيضاً تعطيل الأغشية الحيوية القائمة وقتل الخلايا المقيمة، رغم أن تركيزاً أعلى مطلوب مقارنة بالخلايا العوالقية (Okhiria et al., 2009; Maddocks et al., 2013; Lu et al., 2014; Majtan et al., 2014a). مؤخراً، تم اختبار عسل Manuka على غشاء حيوي متعدد الأنواع يحتوي على Staphylococcus aureus, Streptococcus agalactiae, Pseudomonas aeruginosa، وEnterococcus faecalis ووجد أنه يقلل من حيوية جميع الأنواع باستثناء E. faecalis، التي لم يمكن القضاء عليها (Sojka et al., 2016). وهذا له دلالات سريرية واضحة لاستخدام العسل على الجروح التي تحتوي على أغشية حيوية، وفهم كيف يمكن للغشاء الحيوي أن يمكن E. faecalis من البقاء على قيد الحياة رغم أنه عادةً يُقتل بالعسل هو مجال مهم ومثير للدراسة المستقبلية. يبدو أن MGO مسؤول إلى حد كبير ولكن ليس بالكامل عن تثبيط الأغشية الحيوية بواسطة عسل Manuka، مما يبرز مرة أخرى أهمية المكونات الإضافية التي تعدل النشاط (Kilty et al., 2011; Lu et al., 2014).
لم يتم بعد تحديد نطاق نشاط العسل تجاه مسببات الأمراض غير البكتيرية بشكل جيد. أشارت دراسات حديثة تفحص التأثير المضاد للفيروسات لعسل Manuka إلى أنه يمتلك إمكانات لعلاج فيروس جدري الماء والحزام الناري (السبب في جدري الماء والقوباء المنطقية) (Shahzad and Cohrs, 2012) والإنفلونزا (Watanabe et al., 2014). مسببات الأمراض الفطرية للجلد، بما في ذلك Candida albicans وأنواع الفطريات الجلدية أقل حساسية بكثير من البكتيريا تجاه عسل Manuka، لكنها تُثبط بواسطة العسل الذي ينتج مستويات عالية من بيروكسيد الهيدروجين (Brady et al., 1996; Irish et al., 2006). وُجد أن عسل Manuka وغير Manuka يقلل من حيوية أبواغ الميكروسبوريديان Nosema apis، وهو مسبب مرض مهم للنحل، لكن العسل لم يستطع علاج عدوى النحل بمجرد بدء الإصابة (Malone et al., 2001). هناك عدد قليل جدًا من الدراسات حول استخدام العسل للطفيليات الأولية أو الديدان، ولم تستخدم هذه الدراسات عسلًا ذا نشاط موصوف جيدًا، مما يصعب تقييم أهمية نتائجها (Bassam et al., 1997; Nilforoushzadeh et al., 2007; Sajid and Azim, 2012).
إدخال العسل في الطب السائد: دراسات تجريبية وآلية حديثة تسلط الضوء على كيفية عمل العسل
عسل المانوكا النشط متوفر على نطاق واسع كعامل علاجي وطعام وظيفي، ويقبله معظم المستهلكين كمنتج شامل وغامض إلى حد ما. ومع ذلك، فإن نقص الفهم لكيفية قتل العسل للبكتيريا وتعزيز الشفاء يحد من قبوله في الطب السائد حيث لا يزال يُعتبر "بديلًا" أو "مكملًا". الغالبية العظمى من الدراسات البحثية حول العسل حتى الآن كانت وصفية، لكن الدراسات الحديثة تحاول فك كيفية عمل العسل وتستخدم مناهج ميكانيكية لتحديد كيفية تأثيره على المستوى الخلوي والجزيئي.
دراسات فائقة البنية على الخلايا والمجتمعات البكتيرية المعالجة بالعسل
يمكن للعسل أن يغير بشكل عميق حجم وشكل الخلايا البكتيرية، على الرغم من أن مدى هذا التغيير يختلف بين أنواع البكتيريا المختلفة. باستخدام المجهر الإلكتروني الناقل (TEM)، أظهرت مزروعات S. aureus المعالجة بعسل المانوكا وجود عدد أكبر من الخلايا التي أكملت الانقسام مقارنة بتلك المعالجة بالعسل الصناعي، مما يشير إلى أن الخلايا دخلت مرحلة الانقسام لكنها فشلت في إكمالها، على الرغم من أن هذه الخلايا بدت طبيعية من الخارج عند فحصها بالمجهر الإلكتروني الماسح (SEM) (Henriques et al., 2010). وفي دراسة أحدث، أظهرت الصور بتقنية التباين الطوري بعد المعالجة بجرعة دون مميتة من عسل المانوكا أن خلايا S. aureus وBacillus subtilis كانت أصغر بشكل ملحوظ وأكثر احتمالاً لاحتواء حمض نووي مكثف مقارنة بالخلايا التي نمت بدون عسل (Lu et al., 2013). من الصعب مقارنة هذه الدراسات مباشرة لأنها استخدمت كميات وأوقات معالجة مختلفة من العسل، لكن النتائج بشكل عام تشير إلى انفصال بين النمو والانقسام الخلوي، وهو أمر يُرى غالبًا استجابةً للإجهادات الغذائية والبيئية (Silva-Rocha and de Lorenzo, 2010).
تم الإبلاغ عن أن علاج العسل يسبب في ثقافات الأنواع سالبة الجرام E. coli وP. aeruginosa وجود خلايا أقصر وأطول بشكل غير طبيعي (لو وآخرون، 2013). ومن المثير للاهتمام، بينما يبدو أن P. aeruginosa أقل عرضة للتثبيط بواسطة العسل مقارنة بالأنواع الأخرى، لوحظت تغييرات خلوية عميقة باستخدام المجهر الإلكتروني النافذ والمجهر الإلكتروني الماسح، بما في ذلك الأخاديد والنتوءات (بروزات في أغشية البلازما الخلوية) على سطح الخلية وكمية كبيرة من الحطام خارج الخلية مما يشير إلى تحلل الخلايا (هنريكيس وآخرون، 2011). تم التحقق من ذلك في دراسة لاحقة باستخدام تلوين باكلايت الحي-الميت والتصوير المجهري المرافق، على الرغم من أن ذلك أظهر أيضًا بقاء عدد كبير نسبيًا من الخلايا الحية. استخدمت هذه الدراسات عسلًا بنسبة 20% (وزن/حجم)، وهي نسبة أعلى من الحد الأدنى لتركيز القتل للبكتيريا (MBC) لسلالة P. aeruginosa الخاصة بهم، وكان من المتوقع حدوث تثبيط وموت كبيرين. ومع ذلك، وجد المجهر القوة الذرية (AFM) باستخدام مستويات دون القاتلة للبكتيريا تشوهًا كبيرًا في الخلايا ونتوءات في الخلايا المعالجة بتركيزات MIC (12%) ونصف MIC (6%)، إلى جانب تحلل خلوي كبير (روبرتس وآخرون، 2012). وقد دعم هذا التدهور الظاهر لخلية P. aeruginosa تحليل PCR الكمي الذي أظهر انخفاضًا بمقدار عشرة أضعاف في التعبير عن oprF في الخلايا المعالجة بالعسل، وهو الجين الذي يشفر لبورين الغشاء الخارجي المهم للاستقرار الهيكلي (جينكينز وآخرون، 2015a).
تحليلات الأوميكس تقيم استجابة الخلية الكاملة للتثبيط بواسطة العسل
لقد أحدثت القدرة على تقييم مخرجات الخلية الكاملة ثورة في دراسة تفاعلات الدواء مع الممرضات ولها قيمة خاصة للمنتجات الطبيعية المعقدة مثل العسل حيث من المحتمل أن تؤثر على عمليات متعددة. أشارت دراسات المصفوفات الدقيقة والبروتيوميات للبكتيريا المعرضة للعسل إلى تحفيز عمليات مرتبطة بالإجهاد وكبت تخليق البروتين (بلير وآخرون، 2009; جينكينز وآخرون، 2011; باكر وآخرون، 2012). وعلى الرغم من أن هذا استجابة نموذجية للعوامل المثبطة، إلا أن العسل أنتج "بصمة" فريدة من التعبير التفريقي شملت العديد من البروتينات ذات الوظائف الافتراضية أو غير المعروفة، مما يشير إلى نمط عمل جديد. الجينات أو البروتينات المحددة التي وُجد أنها منخفضة التنظيم في تحليلات الأوميكس لـS. aureus وE. coli O157/H7 لها وظائف تتعلق بالضراوة، واستشعار الكثافة، وتكوين الأغشية الحيوية (لي وآخرون، 2011; جينكينز وآخرون، 2013)، وفي P. aeruginosa كان هناك انخفاض في تنظيم البروتينات المشاركة في التراكيب الهدبية (روبرتس وآخرون، 2015). هذه الظواهر حاسمة للممرضات لإقامة وإحداث عدوى غازية وتشير إلى أنه بالإضافة إلى تثبيط النمو، يمكن للعسل تقليل القدرة المرضية للبكتيريا المسببة للعدوى.
على الرغم من محدودية عدد ونطاق التحليلات الأوميكس التي أجريت حتى الآن، تشير إلى استجابة خلوية معقدة للعسل مع تباين كبير في أنواع البكتيريا المختلفة. هناك حاجة الآن إلى مناهج متقدمة في علم الأحياء النظامي تسمح بوضع البيانات في سياقها، ودراسات تحقق باستخدام PCR الكمي وسلالات حذف الجينات لفك هذا التعقيد، وقد تكشف هذه عن طرق جديدة للعلاجات الدوائية تهدف إلى تثبيط نمو البكتيريا (هدسون وآخرون، 2012).
التفاعلات بين العسل والمضادات الحيوية التقليدية
بالإضافة إلى استخدامه كعامل وحيد، هناك مجال لاستخدام العسل لتعزيز العلاج بالمضادات الحيوية التقليدية. قد يكون لهذا قيمة خاصة عند دمجه مع العوامل الجهازية التي يمكن توصيلها إلى سرير الجرح عبر الدورة الدموية بينما يُطبق العسل موضعيًا. يمكن للعلاجات المجمعة أيضًا تقليل الجرعة العلاجية للعوامل المضادة للميكروبات ومنع تطور المقاومة، وفي بعض الحالات يمكن أن تؤدي إلى تآزر دوائي، حيث يكون النشاط المشترك أكبر من مجموع الأنشطة الفردية لكل دواء.
أظهرت الدراسات في المختبر التي جمعت بين عسل المانوكا المعتمد علاجيًا وعوامل المضادات الحيوية تأثيرًا تآزريًا مع الأوكساسيلين، التتراسيكلين، الإيميبينيم والموبيروسين ضد نمو سلالة MRSA (جينكينز وكوبر، 2012). علاوة على ذلك، أدى وجود تركيز دون المثبط من العسل مع الأوكساسيلين إلى استعادة حساسية سلالة MRSA للأوكساسيلين. وجد المؤلفون تقليل تنظيم mecR1، الذي يشفر بروتين ربط البنسلين الخاص بـ MRSA (PBP2A) واقترحوا ذلك كآلية للتآزر مع العسل. كما وُجد نشاط تآزري قوي بين عسل المانوكا والريفامبيسين ضد عدة سلالات من S. aureus، بما في ذلك العينات السريرية وسلالات MRSA، ومنع وجود العسل ظهور مقاومة الريفامبيسين في المختبر (مولر وآخرون، 2013). وهذا ذو أهمية سريرية لأن الريفامبيسين يخترق الأنسجة والخراجات جيدًا ويستخدم عادة لعلاج العدوى السطحية بالمكورات العنقودية، لكنه يسبب مقاومة بسرعة ولذلك يجب استخدامه مع عامل آخر. ومن النتائج الإضافية لهذه الدراسة أن التآزر لم يكن بسبب MGO، حيث أن العسل الصناعي المضاف إليه MGO لم يظهر تآزرًا مع الريفامبيسين.
فهم كيفية تأثير العسل على عمل المضادات الحيوية ذات آليات العمل المعروفة جيدًا قد يعزز أيضًا فهمنا لكيفية تأثير العسل على مسببات الأمراض البكتيرية. قام ليو وآخرون (2014) بتوسيع تحليل التآزر ليشمل مضادات حيوية إضافية وسلالات مختلفة من S. aureus وMRSA. اقترحوا أن زيادة الحساسية لكليندامايسين وجنتاميسين قد تنتج عن التأثير المشترك لتقليل تخليق البروتين بواسطة العسل مع تثبيط الريبوسومات بواسطة المضادات الحيوية، في حين أن التآزر مع مضادات بيتا-لاكتام قد يكون بسبب زيادة الإجهاد التأكسدي الناتج عن كلا الطرفين. وبما أن سلالات S. aureus وMRSA كانت حساسة بشكل متساوٍ لمزيج الأوكساسيلين والعسل، بدا أن التآزر من غير المحتمل أن يكون بسبب تقليل تنظيم PBP2A. ومع ذلك، في عينة سريرية واحدة من MRSA، لم يكن هناك زيادة في الحساسية لكليندامايسين أو جنتاميسين عند وجود العسل، وهو أمر ملحوظ لأنه أول حالة تم الإبلاغ عنها لاختلاف في الاستجابة للعسل بين MRSA وS. aureus. سيكون من المثير للاهتمام دراسة هذا الاختلاف الخاص بالسلالة باستخدام تحليلات النسخ أو البروتيوم في الأبحاث المستقبلية (ليو وآخرون، 2014).
دليل الفعالية من الدراسات الحيوانية، تقارير الحالات، والتجارب السريرية
تروج الشركات التي تنتج وتسوق عسل المانوكا لمعايير أخلاقية عالية وتثني عن استخدام نماذج الحيوانات لدراسة العدوى وشفاء الجروح. ومع ذلك، تم استخدام عسل المانوكا لعلاج الحيوانات التي تعاني من جروح جراحية أو عرضية، لا سيما الخيول، مع نتائج إيجابية (Dart et al., 2015; Bischofberger et al., 2016). وقد لاحظت تقارير الحالات التي استخدمت العسل للجروح والقرح غير الشافية تحسنًا كبيرًا مع زوال العدوى حيث فشلت المضادات الحيوية التقليدية (Regulski, 2008; Smith et al., 2009). ومع ذلك، على الرغم من ذلك والأدلة من العديد من النماذج in vitro وin vivo التي تثبت أن العسل يقتل مسببات الأمراض المسببة للمشاكل في الجروح، هناك نقص في البيانات السريرية القوية لعسل المانوكا. هناك أسباب مختلفة لذلك، بما في ذلك الصعوبات التقنية في إجراء تجربة مزدوجة التعمية محكومة بالدواء الوهمي على مادة مميزة مثل العسل، والاعتبارات الأخلاقية، وقلة اهتمام الممارسين السريريين، والتكلفة مقابل الفائدة لشركات العسل التي تركز على المنتجات الطبيعية والمبيعات بدون وصفة حيث يفرض عسل المانوكا والضمادات المرتبطة به سعرًا مرتفعًا بالفعل. قد تتغير هذه الأمور مع تآكل مقاومة المضادات الحيوية لخيارات العلاج الحالية والبحوث المستمرة التي تبرز إمكانات العسل وتجذب انتباه الممارسين الطبيين.
الفجوات والفرص الناشئة في دراسة العسل
تم إحراز تقدم كبير مؤخرًا في فهمنا للعسل العلاجي، ومع ذلك يظل استخدامه في الطب السريري محدودًا، حتى عندما تبدأ المضادات الحيوية التقليدية في الفشل. تعقيد العسل، الذي يمكن اعتباره أعظم قوته في قتل مسببات الأمراض المتنوعة ومنع المقاومة، يعقد دراسته حيث من المحتمل أن تؤثر العديد من العوامل التي تعمل معًا على النشاط. نحن ندعو إلى المزيد من الدراسات الميكانيكية باستخدام عسل المانوكا العلاجي المسجل بشكل مناسب، لا سيما الدراسات التي تستخدم مناهج علم الأحياء النظامية غير الاختزالية، إلى جانب تحليلات كيميائية وميكروبيولوجية مفصلة لتوضيح كيفية عمل العسل على المستوى الجزيئي والخلوي والسكاني، وكيف يمكن أن يختلف ذلك في سلالات وأنواع مختلفة من مسببات الأمراض الميكروبية، وكيف يستجيب الخلية المضيفة (الجدول 2). يمكن للمعلومات المستفادة من هذه الدراسات أن توجه العلاج وتنتج البيانات السريرية المطلوبة لإدخال العسل في الطب السائد؛ لم يعد العلاج البديل المستخدم فقط عندما تفشل كل العلاجات الأخرى.
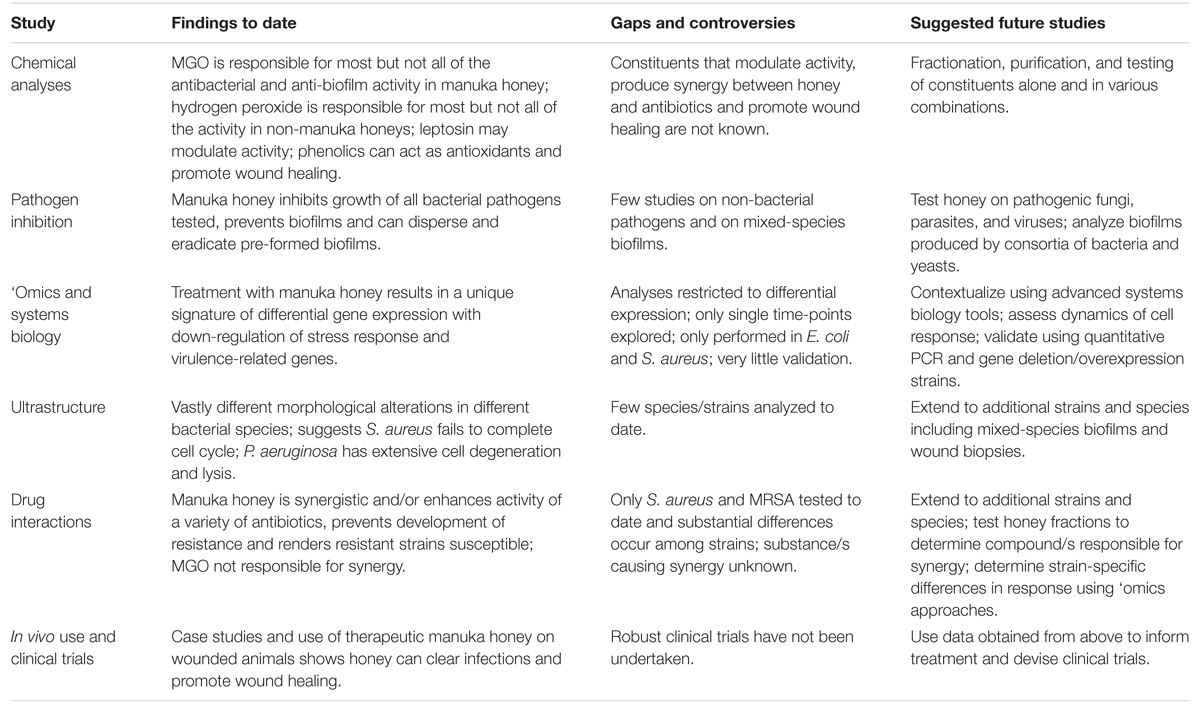
الجدول 2. دراسات عسل المانوكا: النتائج، الفجوات، والدراسات المستقبلية.
مساهمات المؤلفين
تم كتابة هذا الاستعراض بواسطة DC وSB وNNC وDB وPB وتم مراجعته نقديًا بواسطة RS وEH.
التمويل
يحصل NNC على دعم راتب من مؤسسة أبحاث وتطوير الصناعات الريفية – برنامج نحل العسل (المنحة PRJ-009186).
بيان تعارض المصالح
يبلغ DC وPB وEH عن دعم مالي وغير مالي في شكل عسل المانوكا من Comvita NZ Limited وCapilano Honey Limited؛ يعمل RS لدى Comvita NZ Limited، التي تتاجر في عسل المانوكا الطبي (Medihoney).
يعلن بقية المؤلفين أن البحث أُجري في غياب أي علاقة تجارية أو مالية يمكن تفسيرها على أنها تعارض محتمل في المصالح.
الاختصار
ESBL، بيتا-لاكتاماز واسع الطيف؛ MBC، الحد الأدنى لتركيز القاتل للبكتيريا؛ MGO، ميثيل غليوكسال؛ MIC، الحد الأدنى لتركيز التثبيط؛ MRSA، المكورات العنقودية الذهبية المقاومة للميثيسيلين Staphylococcus aureus؛ MRSE، المكورات الجلدية المقاومة للميثيسيلين Staphylococcus epidermis؛ NPA، النشاط غير البيروكسيدي؛ VRE، الإنتيروكوك المقاوم للفانكومايسين Enterococcus.
المراجع
آدامز، س. ج.، بولت، س. هـ.، ديدمان، ب. ج.، فار، ج. م.، غرينجر، م. ن. س.، مانلي-هاريس، م.، وآخرون. (2008). العزل بواسطة HPLC وتوصيف الجزء النشط حيوياً من عسل المانوكا النيوزيلندي (Leptospermum scoparium). بحوث الكربوهيدرات 343، 651–659. doi: 10.1016/j.carres.2007.12.011
آدامز، س. ج.، مانلي-هاريس، م.، ومولان، ب. س. (2009). أصل الميثيلغليوكسال في عسل المانوكا النيوزيلندي (Leptospermum scoparium). بحوث الكربوهيدرات 344، 1050–1053. doi: 10.1016/j.carres.2009.03.020
السمال، ن.، كولي، ك. إ.، مولان، ب. س.، وهانكوك، ب. م. (1994). حساسية Helicobacter pylori للنشاط المضاد للبكتيريا لعسل المانوكا. مجلة الجمعية الملكية للطب 87، 9–12.
ألين، ك.، مولان، ب.، وريد، ج. (1991). مسح للنشاط المضاد للبكتيريا لبعض أنواع العسل في نيوزيلندا. مجلة الصيدلة والصيدلة 43، 817–822. doi: 10.1111/j.2042-7158.1991.tb03186.x
ألين، ك. ل.، ومولان، ب. س. (1997). حساسية البكتيريا المسببة للالتهاب الضرع للنشاط المضاد للبكتيريا للعسل. N. Z. J. Agric. Res. 40، 537–540. doi: 10.1080/00288233.1997.9513276
أنثيميدو، إ.، وموسيالوس، د. (2012). النشاط المضاد للبكتيريا لعسل اليونان وقبرص ضد Staphylococcus aureus وPseudomonas aeruginosa مقارنة بعسل Manuka. J. Med. Food 16، 42–47. doi: 10.1089/jmf.2012.0042
بالان، ب.، مال، ج.، داس، س.، وسينغ، هـ. (2016). الأنشطة المضادة للميكروبات التآزرية والإضافية للكركمين، وعسل Manuka، وبروتينات مصل اللبن. J. Food Biochem. doi: 10.1111/jfbc.12249
بسام، ز.، زهرة، ب. إ.، وسعادة، أ.-أ. (1997). تأثيرات العسل على طفيليات الليشمانيا: دراسة مختبرية. Trop. Doctor27، 36–38.
بيغلاري، ب.، مغدام، أ.، سانتوس، ك.، بليزر، ج.، بوشلر، أ.، يانسن، ج.، وآخرون. (2013). دراسة مستقبلية متعددة المراكز للملاحظة على رعاية الجروح المهنية باستخدام العسل (Medihoney). Int. Wound J. 10، 252–259. doi: 10.1111/j.1742-481X.2012.00970.x
بيشوفبرجر، أ.، دارت، س.، هورادغودا، ن.، بيركنز، ن.، جيفكوت، ل.، ليتل، س.، وآخرون. (2016). تأثير جل Manuka على تركيزات عامل النمو المحول β1 و β3، وعدد البكتيريا، والتشكل النسيجي للجروح الجلدية الملوثة كاملة السماكة في أطراف الخيول البعيدة. Aust. Vet. J. 94، 27–34. doi: 10.1111/avj.12405
Blair, S., Cokcetin, N., Harry, E., و Carter, D. (2009). النشاط المضاد للبكتيريا غير المعتاد لعسل Leptospermum الطبي: طيف النشاط المضاد للبكتيريا، المقاومة، وتحليل النسخ الجيني. Eur. J. Clin. Microbiol. Infect. Dis. 28، 1199–1208. doi: 10.1007/s10096-009-0763-z
Blair, S. E., و Carter, D. A. (2005). الإمكانية العلاجية للعسل في إدارة الجروح والالتهابات. J. Austral. Infect. Control 10، 24–31.
Brady, N., Molan, P., و Harfoot, C. (1996). حساسية الفطريات الجلدية للنشاط المضاد للميكروبات لعسل مانوكا وأنواع العسل الأخرى. Pharm. Pharmacol. Commun. 2، 471–473.
Carnwath, R., Graham, E. M., Reynolds, K., و Pollock, P. J. (2014). النشاط المضاد للميكروبات للعسل ضد عزلات بكتيريا الجروح الشائعة في الخيول. Vet. J. 199، 110–114. doi: 10.1016/j.tvjl.2013.07.003
Chen, C., Campbell, L., Blair, S. E., و Carter, D. A. (2012). تأثير المعالجة الحرارية على الخصائص المضادة للميكروبات في العسل. Front. Microbiol. 3:265. doi: 10.3389/fmicb.2012.00265
Cooper, R., Jenkins, L., Henriques, A., Duggan, R., و Burton, N. (2010). غياب مقاومة البكتيريا لعسل مانوكا الطبي. Eur. J. Clin. Microbiol. Infect. Dis. 29، 1237–1241. doi: 10.1007/s10096-010-0992-1
Cooper, R. A., Halas, E., و Molan, P. C. (2002a). فعالية العسل في تثبيط سلالات Pseudomonas aeruginosa من الحروق المصابة. J. Burn Care Rehabil. 23، 366–370. doi: 10.1097/00004630-200211000-00002
Cooper, R. A., و Molan, P. C. (1999). استخدام العسل كمطهر في إدارة عدوى Pseudomonas. رعاية الجروح 8، 161–164. doi: 10.12968/jowc.1999.8.4.25867
Cooper, R. A., Molan, P. C., و Harding, K. G. (1999). النشاط المضاد للبكتيريا للعسل ضد سلالات Staphylococcus aureus من الجروح المصابة. مجلة الجمعية الملكية للطب. 92، 283–285.
Cooper, R. A., Molan, P. C., و Harding, K. G. (2002b). حساسية المكورات إيجابية الجرام ذات الأهمية السريرية المعزولة من الجروح تجاه العسل. مجلة الميكروبيولوجيا التطبيقية. 93، 857–863. doi: 10.1046/j.1365-2672.2002.01761.x
Cooper, R. A., Wigley, P., و Burton, N. F. (2000). حساسية سلالات Burkholderia cepacia المقاومة المتعددة للعسل. رسائل الميكروبيولوجيا التطبيقية. 31، 20–24. doi: 10.1046/j.1472-765x.2000.00756.x
Dart, A., Bischofberger, A., Dart, C., و Jeffcott, L. (2015). مراجعة للأبحاث حول شفاء جروح الخيول بالنية الثانية باستخدام عسل Manuka: التوصيات الحالية والتطبيقات المستقبلية. تعليم الطب البيطري للخيول. 27، 658–664. doi: 10.1111/eve.12379
Estevinho, L., Pereira, A. P., Moreira, L., Dias, L. G., و Pereira, E. (2008). التأثيرات المضادة للأكسدة والمضادة للميكروبات لمستخلصات المركبات الفينولية من عسل شمال شرق البرتغال. كيمياء الغذاء والسموم. 46، 3774–3779. doi: 10.1016/j.fct.2008.09.062
فرنش، ف. م.، كوبر، ر. أ.، ومولان، ب. س. (2005). النشاط المضاد للبكتيريا للعسل ضد المكورات العنقودية سالبة التخثر. J. Antimicrobial Chemother. 56، 228–231. doi: 10.1093/jac/dki193
جورج، ن. م.، وكاتينغ، ك. ف. (2007). العسل المضاد للبكتيريا (Medihoney): النشاط in-vitro ضد عزلات سريرية من MRSA، VRE، وغيرها من الكائنات سالبة الجرام متعددة المقاومة بما في ذلك Pseudomonas aeruginosa. Wounds 19:231.
هالستيد، ف. د.، ويبّر، م. أ.، راف، م.، بيرت، ر.، درايدن، م.، وأوبنهايم، ب. أ. (2016). النشاط In vitro لعسل مهندَس، وعسل طبي، وضمادات جروح مضادة للميكروبات ضد عزلات بكتيرية سريرية منتجة للغشاء الحيوي. J. Wound Care 25، 93–102. doi: 10.12968/jowc.2016.25.2.93
هاموند، إ. ن.، ودونكور، إ. س. (2013). التأثير المضاد للبكتيريا لعسل Manuka على Clostridium difficile. BMC Res. 6:188. doi: 10.1186/1756-0500-6-188
هنريكيز، أ. ف.، جنكينز، ر. إ.، بيرتون، ن. ف.، وكوبر، ر. أ. (2010). التأثيرات داخل الخلايا لعسل Manuka على Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 29، 45–50. doi: 10.1007/s10096-009-0817-2
هنريكيز، أ. ف.، جنكينز، ر. إ.، بيرتون، ن. ف.، وكوبر، ر. أ. (2011). تأثير عسل Manuka على بنية Pseudomonas aeruginosa. Eur. J. Clin. Microbiol. Infect. Dis. 30، 167–171. doi: 10.1007/s10096-010-1065-1
هادسون، ن. ج.، دالريمبل، ب. ب.، وريفرتر، أ. (2012). ما وراء التعبير التفريقي: البحث عن الطفرات المسببة والجزيئات الفعالة. BMC Genomics 13:356. doi: 10.1186/1471-2164-13-356
آيريش، ج.، بلير، س.، وكارتر، د. (2011). النشاط المضاد للبكتيريا للعسل المستخلص من النباتات الأسترالية. PLoS ONE 6:e18229. doi: 10.1371/journal.pone.0018229
آيريش، ج.، كارتر، د. أ.، شوكوهي، ت.، وبلير، س. إ. (2006). للعسل تأثير مضاد للفطريات ضد أنواع Candida. Med. Mycol. 44، 289–291. doi: 10.1080/13693780600931986
جينكينز، ر.، بيرتون، ن.، وكوبر، ر. (2011). تأثير عسل المانوكا على تعبير بروتين الإجهاد العالمي A في Staphylococcus aureus المقاومة للميثيسيلين. Int. J. Antimicrob. Agents 37، 373–376. doi: 10.1016/j.ijantimicag.2010.11.036
جينكينز، ر.، بيرتون، ن.، وكوبر، ر. (2013). التحليل البروتيني والجيني لـ Staphylococcus aureus المقاومة للميثيسيلين (MRSA) المعرضة لعسل المانوكا in vitro أظهر انخفاضًا في تنظيم علامات الفوعة. J. Antimicrobial Chemother. 69، 603–615. doi: 10.1093/jac/dkt430
جينكينز، ر.، وكوبر، ر. (2012). تحسين فعالية المضادات الحيوية ضد مسببات الأمراض في الجروح باستخدام عسل المانوكا في المختبر. PLoS ONE 7:e45600. doi: 10.1371/journal.pone.0045600
جينكينز، ر.، روبرتس، أ.، وبراون، هـ. ل. (2015أ). حول التأثيرات المضادة للبكتيريا لعسل المانوكا: رؤى ميكانيكية. تقارير أبحاث الأحياء. 6، 215–224. doi: 10.2147/RRB.S75754
جينكينز، ر.، ووتون، م.، هاو، ر.، وكوبر، ر. (2015ب). عرض لقابلية عزلات سريرية مأخوذة من مرضى التليف الكيسي لعسل المانوكا. أرشيف الميكروبيولوجيا. 197، 597–601. doi: 10.1007/s00203-015-1091-6
كالابوس، م. ب. (2008). العلاقة بين الجذور الحرة وميثيلغليوكسال. تفاعلات الكيمياء والبيولوجيا. 171، 251–271. doi: 10.1016/j.cbi.2007.11.009
كاتو، ي.، أوميدا، ن.، مايدا، أ.، ماتسوموتو، د.، كيتاموتو، ن.، وكيكوزاكي، هـ. (2012). تحديد جليكوسيد جديد، ليبتوسين، كعلامة كيميائية لعسل المانوكا. مجلة الكيمياء الزراعية والغذائية. 60، 3418–3423. doi: 10.1021/jf300068w
كيلتي، س. ج.، دوفال، م.، تشان، ف. ت.، فيريس، و.، وسلينجر، ر. (2011). ميثيلغليوكسال: (العامل النشط في عسل المانوكا) النشاط في المختبر ضد الأغشية الحيوية البكتيرية. المنتدى الدولي للحساسية والأنف والجيوب الأنفية. 1، 348–350. doi: 10.1002/alr.20073
كروندا، ج. م.، كوبر، ر. أ.، ومادوكس، س. إ. (2013). عسل المانوكا يثبط إنتاج السايدروفور في Pseudomonas aeruginosa. J. Appl. Microbiol. 115، 86–90. doi: 10.1111/jam.12222
كواكمان، ب. هـ.، تي فيلد، أ. أ.، دي بور، ل.، فاندنبروك-غراولس، س. م.، وزات، س. أ. (2011). لعسلين طبيين رئيسيين آليات مختلفة للنشاط القاتل للبكتيريا. PLoS ONE 6:e17709. doi: 10.1371/journal.pone.0017709
لي، ج.-ه.، بارك، ج.-ه.، كيم، ج.-أ.، نيوبان، ج. ب.، تشو، م. هـ.، لي، س.-س.، وآخرون. (2011). تركيزات منخفضة من العسل تقلل من تكوين الأغشية الحيوية، والتواصل الكمي، والضراوة في Escherichia coli O157: H7. Biofouling 27، 1095–1104. doi: 10.1080/08927014.2011.633704
لين، س. م.، مولان، ب. س.، وكيرسونز، ر. ت. (2011). الحساسية المضبوطة in vitro لمسببات الأمراض المعوية لتأثير عسل المانوكا المضاد للبكتيريا. Eur. J. Clin. Microbiol. Infect. Dis. 30، 569–574. doi: 10.1007/s10096-010-1121-x
ليو، م.، لو، ج.، مولر، ب.، تيرنبل، ل.، بيرك، س. م.، شلوثاور، ر. س.، وآخرون. (2014). اختلافات محددة بالمضادات الحيوية في استجابة Staphylococcus aureus للعلاج بالمضادات الحيوية المدمجة مع عسل المانوكا. Front. Microbiol. 5:779. doi: 10.3389/fmicb.2014.00779
لو، ج., كارتر، د. أ., تيرنبل، ل., روزنديل، د., هيدرلي، د., ستيفنز، ج., وآخرون. (2013). تأثير عسل كانوكا، Manuka، والبرسيم من نيوزيلندا على ديناميكيات نمو البكتيريا وشكل الخلايا يختلف حسب النوع. PLoS ONE 8:e55898. دوى: 10.1371/journal.pone.0055898
لو، ج., تيرنبل، ل., بيرك، س. م., ليو، م., كارتر، د. أ., شلوثاور، ر. س., وآخرون. (2014). أنواع عسل Manuka يمكنها القضاء على الأغشية الحيوية التي تنتجها سلالات Staphylococcus aureus ذات القدرات المختلفة على تكوين الأغشية الحيوية. PeerJ 2:e326. دوى: 10.7717/peerj.326
مادوكز، س. إ., جنكينز، ر. إ., رولاندز، ر. س., بيردي، ك. ج., وكوبر، ر. أ. (2013). عسل Manuka يثبط التصاق وغزو بكتيريا الجروح الطبية المهمة in vitro. مستقبل علم الأحياء الدقيقة 8، 1523–1536. دوى: 10.2217/fmb.13.126
مادوكز، س. إ., لوبيز، م. س., رولاندز، ر. س., وكوبر، ر. أ. (2012). عسل Manuka يثبط تطور الأغشية الحيوية لـStreptococcus pyogenes ويسبب تقليل تعبير بروتينين مرتبطين بالفيبرونيكتين. علم الأحياء الدقيقة 158، 781–790. دوى: 10.1099/mic.0.053959-0
ماجتان، ج. (2014). العسل: منظم مناعي في شفاء الجروح. إصلاح الجروح والتجديد. 22، 187–192. دوى: 10.1111/wrr.12117
ماجتان، ج.، بوهوفا، ج.، هورنياكوفا، م.، كلاوديني، ج.، وماجتان، ف. (2014أ). تأثيرات مضادة لتكوين الأغشية الحيوية للعسل ضد مسببات الأمراض الجرحية Proteus mirabilis وEnterobacter cloacae. Phytother. Res. 28، 69–75. doi: 10.1002/ptr.4957
ماجتان، ج.، بوهوفا، ج.، بروخازكا، إ.، وكلاوديني، ج. (2014ب). قد يؤثر ميثيلغليوكسال على تراكم بيروكسيد الهيدروجين في عسل المانوكا من خلال تثبيط إنزيم جلوكوز أوكسيداز. J. Med. Food 17، 290–293. doi: 10.1089/jmf.2012.0201
ماجتان، ج.، كلاوديني، ج.، بوهوفا، ج.، كوهوتوفا، ل.، دزوروڤا، م.، سيديفا، م.، وآخرون (2012). التعديلات التي يسببها ميثيلغليوكسال على مكونات بروتينية مهمة في نحل العسل في عسل المانوكا: تداعيات علاجية محتملة. Fitoterapia 83، 671–677. doi: 10.1016/j.fitote.2012.02.002
ماجتان، ج.، ماجتانوفا، ل.، بوهوفا، ج.، وماجتان، ف. (2011). عسل الندى كعامل مضاد للبكتيريا قوي في القضاء على عزلات Stenotrophomonas maltophilia المقاومة لعدة أدوية من مرضى السرطان. Phytother. Res. 25، 584–587. doi: 10.1002/ptr.3304
مالون، ل. أ.، غيتهاوس، هـ. س.، وتريجيدغا، إ. ل. (2001). تأثيرات الوقت، درجة الحرارة، والعسل على Nosema apis (Microsporidia: Nosematidae)، طفيلي نحل العسل، Apis mellifera (Hymenoptera: Apidae). J. Invertebrate Pathol. 77، 258–268. doi: 10.1006/jipa.2001.5028
مافريك، E.، ويتتمان، S.، بارت، G.، وهينل، T. (2008). تحديد وقياس الميثيلغليوكسال كمكون مضاد للبكتيريا السائد في عسل Manuka (Leptospermum scoparium) من نيوزيلندا. المغذيات الجزيئية وأبحاث الغذاء 52، 483–489. doi: 10.1002/mnfr.200700282
مولان، P. M. (2008). شرح لماذا مستوى MGO في عسل Manuka لا يظهر النشاط المضاد للبكتيريا. نيوزيلندا بيكيبر 16، 11–13.
مولاي، V.، ومينون، T. (2007). النشاط القاتل للبكتيريا لأنواع مختلفة من العسل ضد عزلات سريرية وبيئية من Pseudomonas aeruginosa. المجلة البديلة للطب التكميلي 13، 439–442. doi: 10.1089/acm.2007.6366
مولر، P.، ألبر، D. G.، تيرنبل، L.، شلوثاور، R. C.، كارتر، D. A.، ويتشرتش، C. B.، وآخرون. (2013). التآزر بين Medihoney والريفامبيسين ضد المكورات العنقودية الذهبية المقاومة للميثيسيلين (MRSA). PLoS ONE 8:e57679. doi: 10.1371/journal.pone.0057679
مندو، M. A.، باديلا-زاكور، O. I.، ووروبو، R. W. (2004). تثبيط نمو مسببات الأمراض المنقولة بالغذاء والكائنات المسببة لتلف الطعام بواسطة أنواع مختارة من العسل الخام. المجلة الدولية لعلم الأحياء الدقيقة الغذائي 97، 1–8. doi: 10.1016/j.ijfoodmicro.2004.03.025
نيلفوروشزاده، م. أ.، جعفري، ف.، مرادي، س.، درخشان، ر.، وهافتبارادران، إ. (2007). تأثير تطبيق العسل الموضعي مع الحقن داخل الآفة للغلوكانتايم في علاج اللشمانيا الجلدية. مجلة الطب التكميلي والبديل BMC 7:1. doi: 10.1186/1472-6882-7-1
نورتون، أ. م.، ماكنزي، ل. ن.، بروكس، ب. ر.، وبابالاردو، ل. ج. (2015). قياس ثنائي هيدروكسي أسيتون في رحيق Leptospermum الأسترالي باستخدام الكروماتوغرافيا السائلة عالية الأداء. مجلة الكيمياء الزراعية والغذائية 63، 6513–6517. doi: 10.1021/acs.jafc.5b01930
أوكيريا، أ.، هينريكيس، أ.، بيرتون، ن.، بيترز، أ.، وكوبر، ر. (2009). العسل يؤثر على الأغشية الحيوية لـ Pseudomonas aeruginosa بطريقة تعتمد على الوقت والجرعة. مجلة منتجات النحل والعلوم الطبية للنحل 1، 6–10. doi: 10.3896/IBRA.4.01.1.03
أوساتو، م. س.، ريدي، س. ج.، وغراهام، د. ي. (1999). التأثير الأسموزي للعسل على نمو وبقاء Helicobacter pylori. Dig. Dis. Sci. 44، 462–464. doi: 10.1023/A:1026676517213
باكر، ج. م.، إيريش، ج.، هربرت، ب. ر.، هيل، س.، بادولا، م.، بلير، س. إ.، وآخرون. (2012). التأثير المضاد للبكتيريا غير المعتمد على البيروكسيد في عسل المانوكا على بروتيوم Staphylococcus aureus. المجلة الدولية للعوامل المضادة للميكروبات 40، 43–50. doi: 10.1016/j.ijantimicag.2012.03.012
روبرتس، أ. إ.، مادوكس، س. إ.، وكوبر، ر. أ. (2012). عسل Manuka قاتل للبكتيريا ضد الزائفة الزنجارية وينتج عنه تعبير تفريقي لجينات oprF و algD. علم الأحياء الدقيقة 158، 3005–3013. دوى: 10.1099/mic.0.062794-0
روبرتس، أ. إ. ل.، مادوكس، س. إ.، وكوبر، ر. أ. (2015). يقلل عسل Manuka من حركة الزائفة الزنجارية عن طريق كبح جينات المرتبطة بالأسواط. مجلة العلاج الكيميائي المضاد للميكروبات 70، 716–725. دوى: 10.1093/jac/dku448
ساجد، م.، وعظيم، م. ك. (2012). توصيف النشاط القاتل للديدان للعسل الطبيعي. مجلة الكيمياء الزراعية والغذائية 60، 7428–7434. دوى: 10.1021/jf301653n
شهزاد، أ.، وكورهس، ر. ج. (2012). النشاط المضاد للفيروسات في المختبر للعسل ضد فيروس جدري الماء (VZV): دراسة طبية ترجميّة لعلاج محتمل للحزام الناري. الطب الحيوي الترجمي 3:2.
شيرلوك، أ.، دولان، أ.، أثمان، ر.، باور، أ.، جيثين، ج.، كومان، س.، وآخرون. (2010). مقارنة النشاط المضاد للميكروبات لعسل أولمو من تشيلي وعسل Manuka ضد المكورات العنقودية الذهبية المقاومة للميثيسيلين، الإشريكية القولونية والزائفة الزنجارية. الطب التكميلي والبديل BMC 10:47. دوى: 10.1186/1472-6882-10-47
سيلفا-روتشا، ر.، ودي لورينزو، ف. (2010). الضوضاء والصلابة في الشبكات التنظيمية بدائية النواة. المراجعة السنوية لعلم الأحياء الدقيقة 64، 257–275. دوى: 10.1146/annurev.micro.091208.073229
سميث، ت., هانفت، ج. ر., وليجيل، ك. (2009). عسل Leptospermum الموضعي في جروح الساق الوريدية المقاومة للعلاج: سلسلة حالات أولية. Adv. Skin Wound Care 22, 68–71. doi: 10.1097/01.ASW.0000345283.05532.9a
سوفر، أ. (1976). كلاب الشيواوا والليتريل، العلاج بالاستخلاب، وعسل من بولدر، كولورادو [editorial]. Arch. Intern. Med.136, 865–866. doi: 10.1001/archinte.136.8.865
سوجكا، م., فالاشوفا، إ., بوتشكوفا، م., وماجتان، ج. (2016). فعالية مضادة للبيوفيلم للعسل والديفنسين-1 المستخلص من النحل على بيوفيلم الجروح متعددة الأنواع. J. Med. Microbiol. doi: 10.1099/jmm.0.000227 [نشر مسبق].
ستيفنز، ج. م., شلوثاور، ر. س., موريس، ب. د., يانغ، د., فيرنلي، ل., غرينوود، د. ر., وآخرون. (2010). المركبات الفينولية والميثيلغليوكسال في بعض أنواع عسل مانوكا وكانوكا من نيوزيلندا. Food Chem. 120, 78–86. doi: 10.1016/j.foodchem.2009.09.074
تان، هـ. ت., رحمن، ر. أ., جان، س. هـ., حليم، أ. س., حسن، س. أ., سليمان، س. أ., وآخرون. (2009). الخصائص المضادة للبكتيريا لعسل توالانج الماليزي ضد الكائنات الدقيقة في الجروح والأمعاء مقارنة بعسل مانوكا. BMC Complement Altern. Med. 9:34. doi: 10.1186/1472-6882-9-34
واتانابي، K.، رحماساري، R.، ماتسوناغا، A.، هاروياما، T.، وكوبايشي، N. (2014). التأثيرات المضادة للفيروسات الإنفلونزا للعسل in vitro: النشاط العالي القوي لعسل manuka honey. Arch. Med. Res. 45، 359–365. doi: 10.1016/j.arcmed.2014.05.006
ويلكينسون، J. M.، وكافانا، H. M. A. (2005). النشاط المضاد للبكتيريا لـ 13 نوعًا من العسل ضد Escherichia coli وPseudomonas aeruginosa. J. Med. Food 8، 100–103. doi: 10.1089/jmf.2005.8.100
ويليامز، S.، كينغ، J.، ريفيل، M.، مانلي-هاريس، M.، بالكز، M.، يانوش، F.، وآخرون. (2014). التغيرات الإقليمية والسنوية والفردية في محتوى الديهيدروكسي أسيتون في رحيق manuka (Leptospermum scoparium) في نيوزيلندا. J. Agric. Food Chem. 62، 10332–10340. doi: 10.1021/jf5045958
ويليكس، D. J.، مولان، P. C.، وهارفوت، C. G. (1992). مقارنة حساسية أنواع البكتيريا المسببة لعدوى الجروح للنشاط المضاد للبكتيريا لعسل manuka وأنواع العسل الأخرى. J. Appl. Bacteriol. 73، 388–394. doi: 10.1111/j.1365-2672.1992.tb04993.x
ويندسور، S.، بابالاردو، M.، بروكس، P.، ويليامز، S.، ومانلي-هاريس، M. (2012). تحليل جديد ملائم للديهيدروكسي أسيتون والميثيلغليوكسال مطبق على عسل Leptospermum الأسترالي. J. Pharmacogn. Phytother. 4، 6–11.
العسل العلاجي من Manuka: لم يعد بديلاً فقط - https://www.frontiersin.org/articles/10.3389/fmicb.2016.00569/full
اترك تعليقًا