Miele di Manuka Terapeutico: Non Più Così Alternativo

La ricerca sul miele medicinale sta vivendo una sostanziale rinascita. Da rimedio popolare largamente ignorato dalla medicina tradizionale come “alternativo”, ora assistiamo a un crescente interesse da parte di scienziati, medici clinici e pubblico generale per gli usi terapeutici del miele. Ci sono diversi fattori che guidano questo interesse: primo, l’aumento della resistenza agli antibiotici di molti patogeni batterici ha stimolato lo sviluppo e l’uso di nuovi antibatterici; secondo, un numero crescente di studi affidabili e casi clinici ha dimostrato che alcuni mieli sono molto efficaci nel trattamento delle ferite; terzo, il miele terapeutico ha un prezzo premium, e l’industria del miele sta attivamente promuovendo studi che le permettano di capitalizzare su questo; infine, la natura molto complessa e piuttosto imprevedibile del miele rappresenta una sfida interessante per gli scienziati di laboratorio. In questo articolo esaminiamo la ricerca sul miele di Manuka, dagli studi osservazionali sui suoi effetti antimicrobici fino ai lavori sperimentali e meccanicistici attuali che mirano a portare il miele nella medicina tradizionale. Delineiamo le lacune attuali e le controversie ancora aperte nella nostra conoscenza di come agisce il miele, e suggeriamo nuovi studi che potrebbero rendere il miele un’alternativa non più “alternativa”.
Introduzione
Il miele è stato usato come medicina nel corso della storia dell'umanità. Uno degli usi terapeutici più comuni e persistenti del miele è stato come medicazione per ferite, quasi certamente grazie alle sue proprietà antimicrobiche. Con l'avvento degli antibiotici altamente attivi negli anni '60, il miele è stato considerato una "sostanza inutile ma innocua" (Soffer, 1976). Tuttavia, la crisi attuale e crescente della resistenza agli antibiotici ha riacceso l'interesse per l'uso del miele, sia come agente efficace di per sé sia come spunto terapeutico per sviluppare nuovi metodi di trattamento. Il miele deriva solitamente dal nettare dei fiori ed è prodotto dalle api, più comunemente dall'ape europea Apis mellifera, ed è un complesso mix di zuccheri, aminoacidi, fenoli e altre sostanze. I tipi di miele derivati da diverse piante da fiore variano sostanzialmente nella loro capacità di uccidere i batteri, e questo ha complicato la letteratura sul miele rendendo talvolta difficile riprodurre i risultati tra studi differenti (Allen et al., 1991; Irish et al., 2011). La maggior parte degli studi recenti che indagano il meccanismo d'azione del miele si sono concentrati sul miele di Manuka attivo, ben caratterizzato e standardizzato, prodotto da alcune specie di Leptospermum native della Nuova Zelanda e dell'Australia, che è stato registrato come prodotto per la cura delle ferite presso gli enti regolatori medici competenti. Pertanto, salvo diversa indicazione, questa recensione si concentrerà sul miele di Manuka.
Analisi chimiche del miele di Manuka attivo
Il professor Peter Molan dell'Università di Waikato, Nuova Zelanda, è stato il primo a segnalare l'attività insolita del miele di Manuka e ha iniziato a testarne l'azione contro un'ampia gamma di specie batteriche a metà degli anni '80. Tuttavia, sebbene fosse chiaro che anche basse concentrazioni di miele di Manuka uccidessero i patogeni batterici, il principio attivo specifico responsabile di ciò è rimasto sfuggente per molti anni. L'elevato contenuto di zucchero e il pH basso rendono il miele inibitorio per la crescita microbica, ma l'attività persiste anche quando questi fattori sono diluiti a livelli trascurabili. Molti diversi tipi di miele producono anche perossido di idrogeno quando la glucosio ossidasi, derivata dall'ape, reagisce con glucosio e acqua. Tuttavia, nel miele di Manuka la produzione di perossido di idrogeno è relativamente bassa e può essere neutralizzata dalla catalasi, eppure l'attività persiste. La causa di questa attività residua, chiamata “attività non perossidica” o NPA, è stata finalmente rivelata nel 2008, quando due laboratori hanno identificato indipendentemente il metilgliossale (MGO) nel miele di Manuka (Adams et al., 2008; Mavric et al., 2008). Il MGO deriva dalla disidratazione spontanea del suo precursore diidrossiacetone (DHA), un fitochimico naturale presente nel nettare dei fiori di Leptospermum scoparium, Leptospermum polygalifolium e di alcune specie correlate di Leptospermum originarie della Nuova Zelanda e dell'Australia (Adams et al., 2009; Williams et al., 2014; Norton et al., 2015). Il MGO può reagire in modo relativamente non specifico con macromolecole come DNA, RNA e proteine (Adams et al., 2008; Mavric et al., 2008; Majtan et al., 2014b) e potrebbe teoricamente essere tossico per le cellule dei mammiferi (Kalapos, 2008). Tuttavia, non ci sono prove di danni alle cellule ospiti quando il miele di Manuka viene consumato per via orale o usato come medicazione per ferite; anzi, il miele sembra stimolare la guarigione e ridurre le cicatrici quando applicato sulle ferite (Biglari et al., 2013; Majtan, 2014; Dart et al., 2015). Come eserciti questa apparente tossicità selettiva verso le cellule batteriche non è noto.
Alti livelli di MGO o perossido di idrogeno solitamente producono il miele più attivo, tuttavia la correlazione non è sempre perfetta, suggerendo che altri componenti del miele possano modulare l'attività (Molan, 2008; Kwakman et al., 2011; Chen et al., 2012; Lu et al., 2013). La defensina-1 delle api, un peptide antimicrobico derivato dalle api, è responsabile dell'attività nel miele Revamil, un miele attivo prodotto da una fonte non divulgata, ma sembra essere strutturalmente modificata e inattiva nel miele Manuka (Kwakman et al., 2011; Majtan et al., 2012). Il livello di leptosina, un glicoside presente esclusivamente nel miele di Leptospermum, è correlato alla potenza e può modulare l'attività antimicrobica del miele Manuka (Kato et al., 2012). Analogamente, vari composti fenolici con potenziale attività antimicrobica possono essere presenti, in particolare nei mieli di colore più scuro, e sebbene si trovino a livelli che da soli difficilmente sono inibitori, possono agire in sinergia tra loro o con altri componenti del miele per produrre o modificare l'attività (Estevinho et al., 2008; Stephens et al., 2010). I fenoli possono anche agire come antiossidanti e potrebbero essere responsabili delle proprietà antinfiammatorie e di guarigione delle ferite del miele (Stephens et al., 2010). Va notato che non tutte le specie di Leptospermum producono miele attivo, e anche all'interno del miele di L. scoparium e L. polygalifolium i livelli di MGO possono variare da ∼100 a >1200 ppm (Windsor et al., 2012). Un'indagine sull'attività del miele australiano ha rilevato che il miele proveniente da piante di Leptospermum che crescono intorno al confine tra New South Wales e Queensland era particolarmente attivo, ma non si sa se ciò sia dovuto a fattori legati alla pianta, al suolo, al clima o ad altri fattori (Irish et al., 2011).
L'inibizione dei patogeni da parte del miele
Il miele è stato testato in vitro su una vasta gamma di patogeni, in particolare quelli che possono colonizzare la pelle, le ferite e le membrane mucose, dove è possibile un trattamento topico con miele. Ad oggi, i test in vitro hanno dimostrato che il miele di Manuka può efficacemente inibire tutti i patogeni batterici problematici testati (riassunti nella Tabella 1). Di particolare interesse è il fatto che gli isolati clinici con fenotipi di resistenza multipla ai farmaci (MDR) non mostrano una riduzione della loro sensibilità al miele, indicando uno spettro d'azione ampio e diverso da qualsiasi antimicrobico conosciuto (Willix et al., 1992; Blair and Carter, 2005; George and Cutting, 2007; Tan et al., 2009). Inoltre, i tentativi di generare ceppi resistenti al miele in laboratorio non hanno avuto successo e non sono stati riportati isolati clinici con resistenza acquisita al miele (Blair et al., 2009; Cooper et al., 2010).
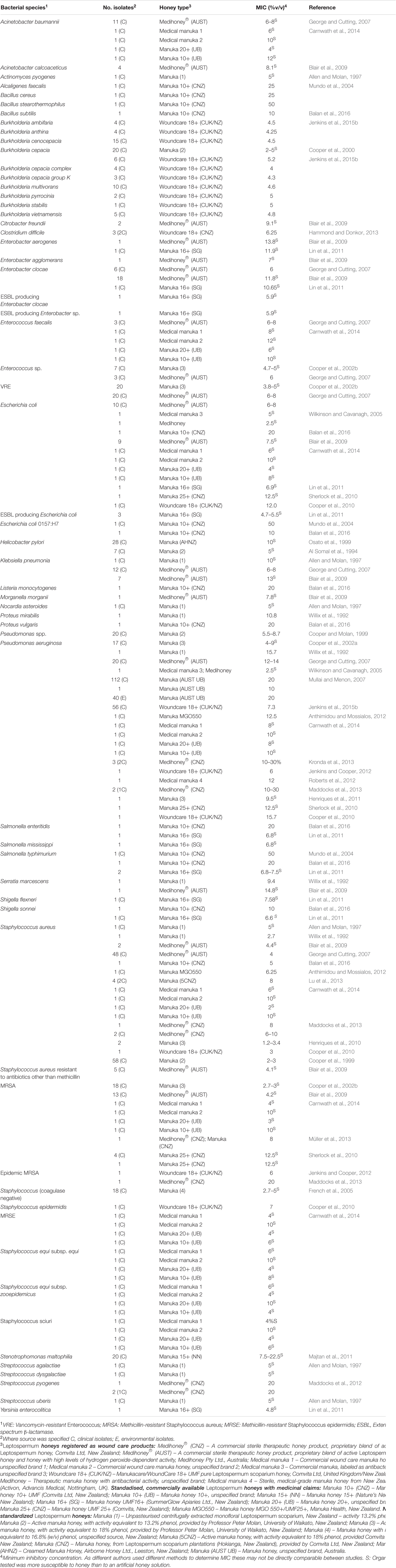
TABELLA 1. Specie batteriche risultate sensibili al miele di Manuka terapeutico.
Oltre a inibire le cellule planctoniche, il miele può disperdere e uccidere i batteri che vivono nei biofilm. I biofilm sono comunità di cellule generalmente racchiuse in una matrice extracellulare autoprodotta e si trovano aderenti a superfici, comprese ferite, denti, superfici mucose e dispositivi impiantati. I microbi residenti nei biofilm sono protetti dagli agenti antimicrobici e possono causare infezioni persistenti e non risolutive. Il miele di Manuka interrompe gli aggregati cellulari (Maddocks et al., 2012; Roberts et al., 2012) e previene la formazione di biofilm da parte di un'ampia gamma di patogeni problematici, inclusi i generi Streptococcus e Staphylococcus, Pseudomonas aeruginosa, Escherichia coli, Proteus mirabilis, Enterobacter cloacae, Acinetobacter baumannii e Klebsiella pneumonia (Maddocks et al., 2012, 2013; Lu et al., 2014; Majtan et al., 2014a; Halstead et al., 2016). È importante sottolineare che il miele può anche interrompere biofilm già stabiliti e uccidere le cellule residenti, sebbene sia necessaria una concentrazione più elevata rispetto alle cellule planctoniche (Okhiria et al., 2009; Maddocks et al., 2013; Lu et al., 2014; Majtan et al., 2014a). Molto recentemente, il miele di Manuka è stato testato su un biofilm multispecie contenente Staphylococcus aureus, Streptococcus agalactiae, Pseudomonas aeruginosa e Enterococcus faecalis ed è stato riscontrato che riduce la vitalità di tutte le specie tranne E. faecalis, che non è stato possibile eradicare (Sojka et al., 2016). Questo ha chiare implicazioni cliniche per l'uso del miele su ferite contenenti biofilm, e comprendere come il biofilm consenta a E. faecalis di sopravvivere quando normalmente viene ucciso dal miele è un'area importante e interessante per studi futuri. Il MGO sembra essere in gran parte, ma non completamente, responsabile dell'inibizione dei biofilm da parte del miele di Manuka, evidenziando ancora una volta l'importanza di componenti aggiuntivi che modulano l'attività (Kilty et al., 2011; Lu et al., 2014).
Lo spettro di attività del miele verso patogeni non batterici deve ancora essere ben definito. Studi recenti che hanno esaminato l'effetto antivirale del miele Manuka hanno suggerito un potenziale trattamento per il virus varicella-zoster (la causa della varicella e dell'herpes zoster) (Shahzad e Cohrs, 2012) e l'influenza (Watanabe et al., 2014). I patogeni fungini della pelle, inclusi Candida albicans e specie di dermatofiti, sono sostanzialmente meno suscettibili rispetto ai batteri al miele Manuka, ma sono inibiti da miele con alti livelli di produzione di perossido di idrogeno (Brady et al., 1996; Irish et al., 2006). È stato riscontrato che miele Manuka e non Manuka riducono la vitalità delle spore del microsporidio Nosema apis, un importante patogeno delle api, ma il miele non è stato in grado di curare l'infezione delle api una volta che questa era in corso (Malone et al., 2001). Ci sono stati pochissimi studi sull'uso del miele per parassiti protozoi o elminti e questi non hanno utilizzato miele con attività ben caratterizzata, rendendo difficile valutare il significato dei loro risultati (Bassam et al., 1997; Nilforoushzadeh et al., 2007; Sajid e Azim, 2012).
Portare il miele nella medicina tradizionale: studi sperimentali e meccanicistici recenti fanno luce su come funziona il miele
Il miele di Manuka attivo è ampiamente disponibile come agente terapeutico e alimento funzionale, e la maggior parte dei consumatori lo accetta come un prodotto olistico e in qualche modo misterioso. Tuttavia, la mancanza di comprensione su come il miele uccida i batteri e favorisca la guarigione limita la sua accettazione dalla medicina tradizionale, dove è ancora considerato “alternativo” o “complementare”. La stragrande maggioranza degli studi sul miele finora è stata descrittiva, tuttavia studi recenti stanno cercando di svelare come funziona il miele e stanno utilizzando approcci meccanicistici per determinare come agisce a livello cellulare e molecolare.
Studi ultrastrutturali delle cellule e comunità batteriche trattate con miele
Il miele può alterare profondamente la dimensione e la forma delle cellule batteriche, anche se l'entità di questo varia tra le diverse specie batteriche. Utilizzando la microscopia elettronica a trasmissione (TEM), le colture di S. aureus trattate con miele di Manuka presentavano più cellule con setti completati rispetto a quelle trattate con miele artificiale, suggerendo che le cellule entrassero ma non completassero la fase di divisione del ciclo cellulare, anche se esternamente queste cellule apparivano normali alla microscopia elettronica a scansione (SEM) (Henriques et al., 2010). Più recentemente, l'imaging a contrasto di fase dopo il trattamento con una dose sub-letale di miele di Manuka ha rilevato che le cellule di S. aureus e Bacillus subtilis erano significativamente più piccole e avevano più probabilità di avere DNA condensato rispetto a quelle cresciute senza miele (Lu et al., 2013). È difficile confrontare direttamente questi studi poiché hanno utilizzato quantità diverse di miele e tempi di trattamento differenti, ma nel complesso i risultati suggeriscono un disaccoppiamento tra crescita e divisione cellulare, spesso osservato in risposta a stress nutrizionali e ambientali (Silva-Rocha and de Lorenzo, 2010).
Il trattamento con miele è stato segnalato come causa di cellule anormalmente più corte e più lunghe nelle colture delle specie Gram negative E. coli e P. aeruginosa (Lu et al., 2013). È interessante notare che, mentre P. aeruginosa sembra essere meno suscettibile all’inibizione da miele rispetto ad altre specie, sono stati osservati profondi cambiamenti cellulari tramite TEM e SEM, inclusi solchi e blebs (protuberanze delle membrane plasmatiche cellulari) sulla superficie cellulare e una quantità sostanziale di detriti extracellulari indicativi di lisi cellulare (Henriques et al., 2011). Ciò è stato confermato in uno studio successivo utilizzando la colorazione fluorescente BacLight live-dead e la microscopia confocale, anche se è stato dimostrato che un numero relativamente elevato di cellule vive rimaneva. Questi studi hanno utilizzato il 20% (p/v) di miele, superiore alla MBC per il loro ceppo di P. aeruginosa, e si sarebbe quindi previsto un’inibizione e una morte sostanziali. Tuttavia, la microscopia a forza atomica (AFM) con livelli sub-battericidi ha comunque rilevato una sostanziale distorsione cellulare e blebbing nelle cellule trattate con concentrazioni MIC (12%) e metà MIC (6%), insieme a una lisi cellulare significativa (Roberts et al., 2012). Questa apparente degenerazione della cellula di P. aeruginosa è stata supportata dall’analisi quantitativa PCR che ha mostrato una regolazione negativa di 10 volte in cellule trattate con miele del gene oprF, che codifica per una porina della membrana esterna importante per la stabilità strutturale (Jenkins et al., 2015a).
Analisi ‘Omiche Valutano la Risposta dell’Intera Cellula all’Inibizione da Parte del Miele
La capacità di valutare le risposte dell’intera cellula ha rivoluzionato lo studio delle interazioni tra farmaci e patogeni ed è particolarmente utile per prodotti naturali complessi come il miele, dove sono probabili effetti su molteplici processi. Studi di microarray e proteomica su batteri esposti al miele hanno suggerito un’induzione di processi legati allo stress e una soppressione della sintesi proteica (Blair et al., 2009; Jenkins et al., 2011; Packer et al., 2012). Sebbene nel complesso questa sia una risposta tipica agli agenti inibitori, il miele ha prodotto una “firma” unica di espressione differenziale che includeva molte proteine con funzioni ipotetiche o sconosciute, suggerendo un nuovo meccanismo d’azione. Specifici geni o proteine risultate down-regolate nelle analisi ‘omiche di S. aureus e E. coli O157/H7 hanno funzioni legate alla virulenza, al quorum sensing e alla formazione di biofilm (Lee et al., 2011; Jenkins et al., 2013), e in P. aeruginosa si è osservata una down-regolazione di proteine coinvolte nella flagellazione (Roberts et al., 2015). Questi fenotipi sono critici per i patogeni per stabilirsi e produrre infezioni invasive e indicano che, oltre a inibire la crescita, il miele può ridurre il potenziale patogeno dei batteri infettanti.
Sebbene ancora relativamente limitate per numero e portata, le analisi ‘omiche condotte finora suggeriscono una risposta cellulare complessa al miele con notevole variazione tra le diverse specie batteriche. Sono ora necessari approcci avanzati di biologia dei sistemi che permettano la contestualizzazione dei dati e studi di validazione mediante PCR quantitativa e ceppi con delezione genica per districare questa complessità, che potrebbero rivelare nuovi approcci per terapie farmacologiche mirate a inibire la crescita batterica (Hudson et al., 2012).
Interazioni tra miele e antibiotici convenzionali
Oltre all'uso come agente unico, esiste la possibilità di utilizzare il miele per integrare il trattamento con antibiotici convenzionali. Questo può avere un valore particolare se combinato con agenti sistemici che possono essere trasportati al letto della ferita tramite la circolazione sanguigna mentre il miele viene applicato localmente. I trattamenti combinati possono anche ridurre la dose terapeutica degli agenti antimicrobici e prevenire lo sviluppo di resistenza, e in alcuni casi possono portare a una sinergia farmacologica, in cui l'attività combinata è maggiore della somma delle attività individuali di ciascun farmaco.
Studi in vitro che combinano miele di manuka terapeuticamente approvato con agenti antibiotici hanno riscontrato un effetto sinergico con oxacillina, tetraciclina, imipenem e mupirocina contro la crescita di un ceppo MRSA (Jenkins e Cooper, 2012). Inoltre, la presenza di una concentrazione sub-inibitoria di miele in combinazione con oxacillina ha ripristinato la suscettibilità del ceppo MRSA all'oxacillina. Gli autori hanno riscontrato una down-regolazione di mecR1, che codifica per una proteina legante la penicillina specifica per MRSA (PBP2A), suggerendo questo come meccanismo della sinergia del miele. È stata inoltre riscontrata una forte attività sinergica tra miele di manuka e rifampicina contro molteplici ceppi di S. aureus, inclusi isolati clinici e ceppi MRSA, e la presenza di miele ha impedito l'emergere di resistenza alla rifampicina in vitro (Müller et al., 2013). Ciò ha rilevanza clinica poiché la rifampicina penetra bene nei tessuti e negli ascessi ed è comunemente usata per trattare infezioni stafilococciche superficiali, ma induce rapidamente resistenza e deve quindi essere usata in combinazione con un altro agente. Un ulteriore risultato di questo studio è stato che la sinergia non era dovuta a MGO, poiché un miele sintetico arricchito con MGO non mostrava sinergia con la rifampicina.
Comprendere come il miele influenzi l'azione degli antimicrobici con modalità d'azione ben caratterizzate può anche approfondire la nostra comprensione di come il miele agisca sui patogeni batterici. Liu et al. (2014) hanno esteso l'analisi della sinergia includendo ulteriori antibiotici e diversi ceppi di S. aureus e MRSA. Hanno suggerito che una maggiore suscettibilità a clindamicina e gentamicina potrebbe derivare dall'effetto combinato della sintesi proteica down-regolata dal miele con l'inibizione dei ribosomi da parte degli antibiotici, mentre la sinergia con gli antibiotici β-lattamici potrebbe essere dovuta all'aumento dello stress ossidativo causato da entrambi i partner. Poiché i ceppi di S. aureus e MRSA erano ugualmente suscettibili alla combinazione oxacillina-miele, sembrava improbabile che la sinergia fosse dovuta alla down-regolazione di PBP2A. Tuttavia, in un isolato clinico di MRSA, non si è osservato un aumento della sensibilità a clindamicina o gentamicina in presenza di miele, il che è notevole in quanto è il primo caso riportato di una differenza nella risposta al miele tra MRSA e S. aureus. Indagare questa differenza specifica di ceppo mediante analisi trascrittomiche o proteomiche sarebbe una interessante direzione per future ricerche (Liu et al., 2014).
Prove di efficacia da studi su animali, casi clinici e trial clinici
Le aziende che producono e commercializzano miele di Manuka promuovono elevati standard etici e scoraggiano l’uso di modelli animali per studiare infezioni e guarigione delle ferite. Tuttavia, il miele di Manuka è stato usato per trattare animali con ferite chirurgiche o accidentali, in particolare cavalli, con risultati positivi (Dart et al., 2015; Bischofberger et al., 2016). Rapporti di casi che utilizzano il miele per ferite e ulcere non guaribili hanno osservato un miglioramento significativo con risoluzione dell’infezione dove gli antibiotici convenzionali avevano fallito (Regulski, 2008; Smith et al., 2009). Tuttavia, nonostante ciò e le evidenze da numerosi modelli in vitro e in vivo che il miele uccide patogeni problematici delle ferite, vi è una scarsità di dati clinici solidi per il miele di Manuka. Ci sono varie ragioni per questo, tra cui difficoltà tecniche nel condurre uno studio controllato con placebo in doppio cieco su una sostanza distintiva come il miele, considerazioni etiche, mancanza di interesse da parte dei medici clinici e rapporto costi-benefici per le aziende del miele, il cui focus è sui prodotti naturali e sulle vendite da banco dove il miele di Manuka e le medicazioni associate già hanno un prezzo premium. Questi fattori potrebbero cambiare man mano che la resistenza agli antibiotici erode le opzioni di trattamento attuali e la ricerca continua che evidenzia il potenziale del miele lo porta all’attenzione dei medici.
Lacune e opportunità emergenti nello studio del miele
Sono stati fatti grandi progressi recentemente nella nostra comprensione del miele terapeutico, eppure il suo uso nella medicina clinica rimane limitato, anche quando gli antibiotici convenzionali cominciano a fallire. La complessità del miele, che è probabilmente la sua più grande forza nel uccidere patogeni diversi e prevenire la resistenza, complica il suo studio poiché molti fattori che agiscono insieme probabilmente influenzano l’attività. Sosteniamo ulteriori studi meccanicistici utilizzando miele terapeutico di Manuka opportunamente registrato, in particolare studi che utilizzano approcci di biologia dei sistemi non riduzionisti, insieme ad analisi chimiche e microbiologiche dettagliate per chiarire come il miele agisce a livello molecolare, cellulare e di popolazione, come questo possa differire in diversi ceppi e specie di patogeni microbici, e come risponde la cellula ospite (Tabella 2). Le informazioni ottenute da questi studi possono quindi informare la terapia e produrre i dati clinici necessari per portare il miele nella medicina tradizionale; non più come terapia alternativa usata solo quando tutto il resto ha fallito.
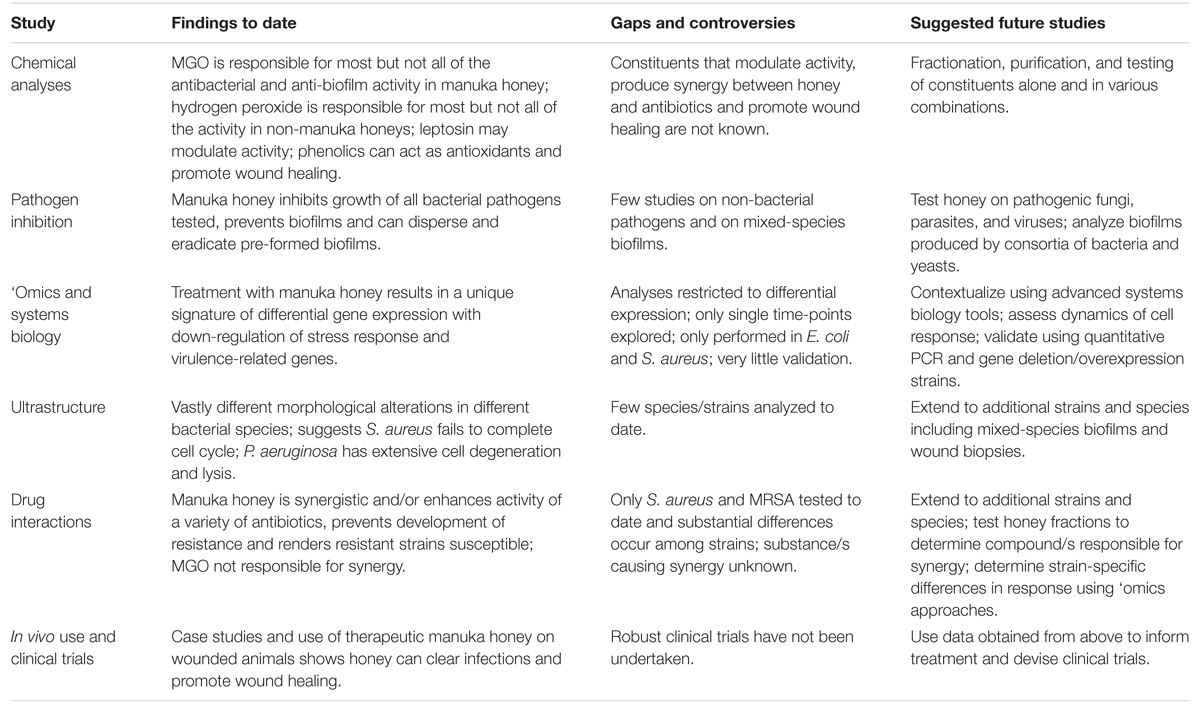
TABELLA 2. Studi sul miele di Manuka: risultati, lacune e studi futuri.
Contributi degli autori
Questa recensione è stata scritta da DC, SB, NNC, DB e PB ed è stata criticamente revisionata da RS ed EH.
Finanziamento
NNC riceve supporto salariale dalla Rural Industries Research and Development Corporation – Honey Bee Program (Grant PRJ-009186).
Dichiarazione di conflitto di interessi
DC, PB e EH riportano supporto finanziario e non finanziario sotto forma di miele di manuka da Comvita NZ Limited e Capilano Honey Limited; RS è impiegato da Comvita NZ Limited, che commercia miele di manuka di grado medico (Medihoney).
Gli altri autori dichiarano che la ricerca è stata condotta in assenza di qualsiasi relazione commerciale o finanziaria che possa essere interpretata come un potenziale conflitto di interessi.
Abbreviazione
ESBL, β-lattamasi a spettro esteso; MBC, concentrazione battericida minima; MGO, metilgliossale; MIC, concentrazione inibitoria minima; MRSA, Staphylococcus aureus resistente alla meticillina; MRSE, Staphylococcus epidermis resistente alla meticillina; NPA, attività non perossidica; VRE, Enterococcus resistente alla vancomicina.
Riferimenti
Adams, C. J., Boult, C. H., Deadman, B. J., Farr, J. M., Grainger, M. N. C., Manley-Harris, M., et al. (2008). Isolamento tramite HPLC e caratterizzazione della frazione bioattiva del miele di manuka (Leptospermum scoparium) della Nuova Zelanda. Carbohydr. Res. 343, 651–659. doi: 10.1016/j.carres.2007.12.011
Adams, C. J., Manley-Harris, M., e Molan, P. C. (2009). L'origine del metilgliossale nel miele di manuka (Leptospermum scoparium) della Nuova Zelanda. Carbohydr. Res. 344, 1050–1053. doi: 10.1016/j.carres.2009.03.020
Al Somal, N., Coley, K. E., Molan, P. C., e Hancock, B. M. (1994). Suscettibilità di Helicobacter pylori all'attività antibatterica del miele di manuka. J. R. Soc. Med. 87, 9–12.
Allen, K., Molan, P., e Reid, G. (1991). Un'indagine sull'attività antibatterica di alcuni mieli della Nuova Zelanda. J. Pharm. Pharmacol.43, 817–822. doi: 10.1111/j.2042-7158.1991.tb03186.x
Allen, K. L., e Molan, P. C. (1997). La sensibilità dei batteri responsabili della mastite all'attività antibatterica del miele. N. Z. J. Agric. Res. 40, 537–540. doi: 10.1080/00288233.1997.9513276
Anthimidou, E., e Mossialos, D. (2012). Attività antibatterica dei mieli greci e ciprioti contro Staphylococcus aureus e Pseudomonas aeruginosa in confronto al Manuka honey. J. Med. Food 16, 42–47. doi: 10.1089/jmf.2012.0042
Balan, P., Mal, G., Das, S., e Singh, H. (2016). Attività antimicrobiche sinergiche e additive di curcumina, Manuka honey e proteine del siero. J. Food Biochem. doi: 10.1111/jfbc.12249
Bassam, Z., Zohra, B. I., e Saada, A.-A. (1997). Gli effetti del miele su Leishmania parasites: uno studio in vitro. Trop. Doctor27, 36–38.
Biglari, B., Moghaddam, A., Santos, K., Blaser, G., Büchler, A., Jansen, G., et al. (2013). Studio osservazionale prospettico multicentrico sulla cura professionale delle ferite con miele (Medihoney). Int. Wound J. 10, 252–259. doi: 10.1111/j.1742-481X.2012.00970.x
Bischofberger, A., Dart, C., Horadagoda, N., Perkins, N., Jeffcott, L., Little, C., et al. (2016). Effetto del gel di Manuka honey sulle concentrazioni di transforming growth factor β1 e β3, sul conteggio batterico e sull'istomorfologia di ferite cutanee contaminate a spessore totale negli arti distali equini. Aust. Vet. J. 94, 27–34. doi: 10.1111/avj.12405
Blair, S., Cokcetin, N., Harry, E., e Carter, D. (2009). L'insolita attività antibatterica del miele di Leptospermum di grado medico: spettro antibatterico, resistenza e analisi del trascrittoma. Eur. J. Clin. Microbiol. Infect. Dis. 28, 1199–1208. doi: 10.1007/s10096-009-0763-z
Blair, S. E., e Carter, D. A. (2005). Il potenziale del miele nella gestione di ferite e infezioni. J. Austral. Infect. Control 10, 24–31.
Brady, N., Molan, P., e Harfoot, C. (1996). La sensibilità dei dermatofiti all'attività antimicrobica del miele di Manuka e di altri mieli. Pharm. Pharmacol. Commun. 2, 471–473.
Carnwath, R., Graham, E. M., Reynolds, K., e Pollock, P. J. (2014). L'attività antimicrobica del miele contro isolati batterici comuni di ferite equine. Vet. J. 199, 110–114. doi: 10.1016/j.tvjl.2013.07.003
Chen, C., Campbell, L., Blair, S. E., e Carter, D. A. (2012). L'effetto del trattamento termico sulle proprietà antimicrobiche del miele. Front. Microbiol. 3:265. doi: 10.3389/fmicb.2012.00265
Cooper, R., Jenkins, L., Henriques, A., Duggan, R., e Burton, N. (2010). Assenza di resistenza batterica al miele di Manuka di grado medico. Eur. J. Clin. Microbiol. Infect. Dis. 29, 1237–1241. doi: 10.1007/s10096-010-0992-1
Cooper, R. A., Halas, E., e Molan, P. C. (2002a). L'efficacia del miele nell'inibire ceppi di Pseudomonas aeruginosa da ustioni infette. J. Burn Care Rehabil. 23, 366–370. doi: 10.1097/00004630-200211000-00002
Cooper, R. A., e Molan, P. C. (1999). L’uso del miele come antisettico nella gestione dell’infezione da Pseudomonas. J. Wound Care 8, 161–164. doi: 10.12968/jowc.1999.8.4.25867
Cooper, R. A., Molan, P. C., e Harding, K. G. (1999). Attività antibatterica del miele contro ceppi di Staphylococcus aureusda ferite infette. J. R. Soc. Med. 92, 283–285.
Cooper, R. A., Molan, P. C., e Harding, K. G. (2002b). La sensibilità al miele dei cocchi Gram-positivi di rilevanza clinica isolati da ferite. J. Appl. Microbiol. 93, 857–863. doi: 10.1046/j.1365-2672.2002.01761.x
Cooper, R. A., Wigley, P., e Burton, N. F. (2000). Suscettibilità di ceppi multiresistenti di Burkholderia cepacia al miele. Lett. Appl. Microbiol. 31, 20–24. doi: 10.1046/j.1472-765x.2000.00756.x
Dart, A., Bischofberger, A., Dart, C., e Jeffcott, L. (2015). Una revisione della ricerca sulla guarigione delle ferite equine a seconda intenzione usando miele di Manuka: raccomandazioni attuali e applicazioni future. Equine Vet. Educ. 27, 658–664. doi: 10.1111/eve.12379
Estevinho, L., Pereira, A. P., Moreira, L., Dias, L. G., e Pereira, E. (2008). Effetti antiossidanti e antimicrobici degli estratti di composti fenolici del miele del Nordest del Portogallo. Food Chem. Toxicol. 46, 3774–3779. doi: 10.1016/j.fct.2008.09.062
French, V. M., Cooper, R. A., e Molan, P. C. (2005). L'attività antibatterica del miele contro Stafilococchi coagulasi-negativi. J. Antimicrobial Chemother. 56, 228–231. doi: 10.1093/jac/dki193
George, N. M., e Cutting, K. F. (2007). Miele antibatterico (Medihoney): attività in vitro contro isolati clinici di MRSA, VRE e altri organismi gram-negativi multiresistenti inclusi Pseudomonas aeruginosa. Wounds 19:231.
Halstead, F. D., Webber, M. A., Rauf, M., Burt, R., Dryden, M., e Oppenheim, B. A. (2016). Attività in vitro di un miele ingegnerizzato, mieli di grado medico e medicazioni antimicrobiche per ferite contro isolati clinici batterici produttori di biofilm. J. Wound Care 25, 93–102. doi: 10.12968/jowc.2016.25.2.93
Hammond, E. N., e Donkor, E. S. (2013). Effetto antibatterico del miele di Manuka su Clostridium difficile. BMC Res. 6:188. doi: 10.1186/1756-0500-6-188
Henriques, A. F., Jenkins, R. E., Burton, N. F., e Cooper, R. A. (2010). Gli effetti intracellulari del miele di Manuka su Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 29, 45–50. doi: 10.1007/s10096-009-0817-2
Henriques, A. F., Jenkins, R. E., Burton, N. F., e Cooper, R. A. (2011). L'effetto del miele di Manuka sulla struttura di Pseudomonas aeruginosa. Eur. J. Clin. Microbiol. Infect. Dis. 30, 167–171. doi: 10.1007/s10096-010-1065-1
Hudson, N. J., Dalrymple, B. P., e Reverter, A. (2012). Oltre l'espressione differenziale: la ricerca di mutazioni causali e molecole effettrici. BMC Genomics 13:356. doi: 10.1186/1471-2164-13-356
Irish, J., Blair, S., e Carter, D. (2011). L'attività antibatterica del miele derivato dalla flora australiana. PLoS ONE 6:e18229. doi: 10.1371/journal.pone.0018229
Irish, J., Carter, D. A., Shokohi, T., e Blair, S. E. (2006). Il miele ha un effetto antifungino contro specie di Candida. Med. Mycol. 44, 289–291. doi: 10.1080/13693780600931986
Jenkins, R., Burton, N., e Cooper, R. (2011). Effetto del miele di Manuka sull'espressione della proteina universale di stress A in Staphylococcus aureus meticillino-resistente. Int. J. Antimicrob. Agents 37, 373–376. doi: 10.1016/j.ijantimicag.2010.11.036
Jenkins, R., Burton, N., e Cooper, R. (2013). Analisi proteomica e genomica di Staphylococcus aureus meticillino-resistente (MRSA) esposto a miele di Manuka in vitro ha dimostrato una down-regolazione dei marcatori di virulenza. J. Antimicrobial Chemother. 69, 603–615. doi: 10.1093/jac/dkt430
Jenkins, R., e Cooper, R. (2012). Migliorare l'attività antibiotica contro i patogeni delle ferite con miele di Manuka in vitro. PLoS ONE 7:e45600. doi: 10.1371/journal.pone.0045600
Jenkins, R., Roberts, A., e Brown, H. L. (2015a). Sugli effetti antibatterici del miele di Manuka: approfondimenti meccanicistici. Res. Rep. Biol. 6, 215–224. doi: 10.2147/RRB.S75754
Jenkins, R., Wootton, M., Howe, R., e Cooper, R. (2015b). Dimostrazione della suscettibilità degli isolati clinici ottenuti da pazienti con fibrosi cistica al miele di Manuka. Arch. Microbiol. 197, 597–601. doi: 10.1007/s00203-015-1091-6
Kalapos, M. P. (2008). Il tandem dei radicali liberi e del metilgliossale. Chem. Biol. Interact. 171, 251–271. doi: 10.1016/j.cbi.2007.11.009
Kato, Y., Umeda, N., Maeda, A., Matsumoto, D., Kitamoto, N., e Kikuzaki, H. (2012). Identificazione di un nuovo glicoside, leptosina, come marcatore chimico del miele di Manuka. J. Agric. Food Chem. 60, 3418–3423. doi: 10.1021/jf300068w
Kilty, S. J., Duval, M., Chan, F. T., Ferris, W., e Slinger, R. (2011). Metilgliossale: (agente attivo del miele di Manuka) attività in vitro contro i biofilm batterici. Int. Forum Allergy Rhinol. 1, 348–350. doi: 10.1002/alr.20073
Kronda, J. M., Cooper, R. A., e Maddocks, S. E. (2013). Il miele di Manuka inibisce la produzione di siderofori in Pseudomonas aeruginosa. J. Appl. Microbiol. 115, 86–90. doi: 10.1111/jam.12222
Kwakman, P. H., te Velde, A. A., de Boer, L., Vandenbroucke-Grauls, C. M., e Zaat, S. A. (2011). Due mieli medicinali principali hanno meccanismi diversi di attività battericida. PLoS ONE 6:e17709. doi: 10.1371/journal.pone.0017709
Lee, J.-H., Park, J.-H., Kim, J.-A., Neupane, G. P., Cho, M. H., Lee, C.-S., et al. (2011). Basse concentrazioni di miele riducono la formazione di biofilm, il quorum sensing e la virulenza in Escherichia coli O157: H7. Biofouling 27, 1095–1104. doi: 10.1080/08927014.2011.633704
Lin, S. M., Molan, P. C., e Cursons, R. T. (2011). La suscettibilità controllata in vitro dei patogeni gastrointestinali all'effetto antibatterico del miele di Manuka. Eur. J. Clin. Microbiol. Infect. Dis. 30, 569–574. doi: 10.1007/s10096-010-1121-x
Liu, M., Lu, J., Müller, P., Turnbull, L., Burke, C. M., Schlothauer, R. C., et al. (2014). Differenze specifiche agli antibiotici nella risposta di Staphylococcus aureus al trattamento con antimicrobici combinati con miele di Manuka. Front. Microbiol. 5:779. doi: 10.3389/fmicb.2014.00779
Lu, J., Carter, D. A., Turnbull, L., Rosendale, D., Hedderley, D., Stephens, J., et al. (2013). L’effetto dei mieli di kanuka, manuka e trifoglio della Nuova Zelanda sulla dinamica di crescita batterica e sulla morfologia cellulare varia a seconda della specie. PLoS ONE8:e55898. doi: 10.1371/journal.pone.0055898
Lu, J., Turnbull, L., Burke, C. M., Liu, M., Carter, D. A., Schlothauer, R. C., et al. (2014). I mieli di tipo Manuka possono eradicare i biofilm prodotti da ceppi di Staphylococcus aureus con diverse capacità di formazione del biofilm. PeerJ 2:e326. doi: 10.7717/peerj.326
Maddocks, S. E., Jenkins, R. E., Rowlands, R. S., Purdy, K. J., e Cooper, R. A. (2013). Il miele di Manuka inibisce l’adesione e l’invasione di batteri importanti per le ferite in vitro. Fut. Microbiol. 8, 1523–1536. doi: 10.2217/fmb.13.126
Maddocks, S. E., Lopez, M. S., Rowlands, R. S., e Cooper, R. A. (2012). Il miele di Manuka inibisce lo sviluppo di biofilm di Streptococcus pyogenes e causa una ridotta espressione di due proteine leganti la fibronectina. Microbiology 158, 781–790. doi: 10.1099/mic.0.053959-0
Majtan, J. (2014). Miele: un immunomodulatore nella guarigione delle ferite. Wound Repair Regenerat. 22, 187–192. doi: 10.1111/wrr.12117
Majtan, J., Bohova, J., Horniackova, M., Klaudiny, J., e Majtan, V. (2014a). Effetti antibiofilm del miele contro i patogeni delle ferite Proteus mirabilis e Enterobacter cloacae. Phytother. Res. 28, 69–75. doi: 10.1002/ptr.4957
Majtan, J., Bohova, J., Prochazka, E., e Klaudiny, J. (2014b). Il metilgliossale può influenzare l'accumulo di perossido di idrogeno nel miele di Manuka attraverso l'inibizione della glucosio ossidasi. J. Med. Food 17, 290–293. doi: 10.1089/jmf.2012.0201
Majtan, J., Klaudiny, J., Bohova, J., Kohutova, L., Dzurova, M., Sediva, M., et al. (2012). Modifiche indotte dal metilgliossale di componenti proteici significativi dell'ape mellifera nel miele di Manuka: possibili implicazioni terapeutiche. Fitoterapia 83, 671–677. doi: 10.1016/j.fitote.2012.02.002
Majtan, J., Majtanova, L., Bohova, J., e Majtan, V. (2011). Miele di melata come potente agente antibatterico nell'eradicazione di isolati multi-resistenti di Stenotrophomonas maltophilia da pazienti oncologici. Phytother. Res. 25, 584–587. doi: 10.1002/ptr.3304
Malone, L. A., Gatehouse, H. S., e Tregidga, E. L. (2001). Effetti del tempo, della temperatura e del miele su Nosema apis (Microsporidia: Nosematidae), un parassita dell'ape mellifera, Apis mellifera (Hymenoptera: Apidae). J. Invertebrate Pathol. 77, 258–268. doi: 10.1006/jipa.2001.5028
Mavric, E., Wittmann, S., Barth, G., e Henle, T. (2008). Identificazione e quantificazione del metilgliossale come principale componente antibatterico dei mieli di Manuka (Leptospermum scoparium) della Nuova Zelanda. Mol. Nutrit. Food Res. 52, 483–489. doi: 10.1002/mnfr.200700282
Molan, P. M. (2008). Una spiegazione del motivo per cui il livello di MGO nel miele di Manuka non mostra l’attività antibatterica. New Zealand Beekeeper 16, 11–13.
Mullai, V., e Menon, T. (2007). Attività battericida di diversi tipi di miele contro isolati clinici e ambientali di Pseudomonas aeruginosa. J. Alternat. Complement. Med. 13, 439–442. doi: 10.1089/acm.2007.6366
Müller, P., Alber, D. G., Turnbull, L., Schlothauer, R. C., Carter, D. A., Whitchurch, C. B., et al. (2013). Sinergia tra Medihoney e rifampicina contro Staphylococcus aureus meticillino-resistente (MRSA). PLoS ONE 8:e57679. doi: 10.1371/journal.pone.0057679
Mundo, M. A., Padilla-Zakour, O. I., e Worobo, R. W. (2004). Inibizione della crescita di patogeni alimentari e organismi di deterioramento alimentare da parte di alcuni mieli grezzi. Int. J. Food Microbiol. 97, 1–8. doi: 10.1016/j.ijfoodmicro.2004.03.025
Nilforoushzadeh, M. A., Jaffary, F., Moradi, S., Derakhshan, R., e Haftbaradaran, E. (2007). Effetto dell’applicazione topica di miele insieme all’iniezione intralesionale di glucantime nel trattamento della leishmaniosi cutanea. BMC Complement Altern. Med. 7:1. doi: 10.1186/1472-6882-7-1
Norton, A. M., McKenzie, L. N., Brooks, P. R., e Pappalardo, L. J. (2015). Quantificazione della diidrossiacetone nel nettare australiano di Leptospermum tramite cromatografia liquida ad alte prestazioni. J. Agric. Food Chem. 63, 6513–6517. doi: 10.1021/acs.jafc.5b01930
Okhiria, O., Henriques, A., Burton, N., Peters, A., e Cooper, R. (2009). Il miele modula i biofilm di Pseudomonas aeruginosa in modo dipendente da tempo e dose. J. ApiProduct. ApiMedical Sci. 1, 6–10. doi: 10.3896/IBRA.4.01.1.03
Osato, M. S., Reddy, S. G., e Graham, D. Y. (1999). Effetto osmotico del miele sulla crescita e vitalità di Helicobacter pylori. Dig. Dis. Sci. 44, 462–464. doi: 10.1023/A:1026676517213
Packer, J. M., Irish, J., Herbert, B. R., Hill, C., Padula, M., Blair, S. E., et al. (2012). Effetto antibatterico specifico non perossidico del miele di Manuka sul proteoma di Staphylococcus aureus. Int. J. Antimicrob. Agents 40, 43–50. doi: 10.1016/j.ijantimicag.2012.03.012
Regulski, M. (2008). Un nuovo medicazione per la cura delle ulcere croniche alle gambe. Podiatry Manag. 27, 235–246.
Roberts, A. E., Maddocks, S. E., e Cooper, R. A. (2012). Il miele di Manuka è battericida contro Pseudomonas aeruginosa e determina un'espressione differenziale di oprF e algD. Microbiology 158, 3005–3013. doi: 10.1099/mic.0.062794-0
Roberts, A. E. L., Maddocks, S. E., e Cooper, R. A. (2015). Il miele di Manuka riduce la motilità di Pseudomonas aeruginosa sopprimendo i geni associati alle flagelle. J. Antimicrob. Chemother. 70, 716–725. doi: 10.1093/jac/dku448
Sajid, M., e Azim, M. K. (2012). Caratterizzazione dell'attività nematocida del miele naturale. J. Agric. Food Chem. 60, 7428–7434. doi: 10.1021/jf301653n
Shahzad, A., e Cohrs, R. J. (2012). Attività antivirale in vitro del miele contro il virus varicella zoster (VZV): uno studio di medicina traslazionale per un potenziale rimedio per l'herpes zoster. Transl. Biomed. 3:2.
Sherlock, O., Dolan, A., Athman, R., Power, A., Gethin, G., Cowman, S., et al. (2010). Confronto dell'attività antimicrobica del miele di Ulmo dal Cile e del miele di Manuka contro Staphylococcus aureus meticillino-resistente, Escherichia coli e Pseudomonas aeruginosa. BMC Complement Altern. Med. 10:47. doi: 10.1186/1472-6882-10-47
Silva-Rocha, R., e de Lorenzo, V. (2010). Rumore e robustezza nelle reti regolatorie procariotiche. Annu. Rev. Microbiol. 64, 257–275. doi: 10.1146/annurev.micro.091208.073229
Smith, T., Hanft, J. R., e Legel, K. (2009). Miele topico di Leptospermum in ferite venose recalcitranti: una serie preliminare di casi. Adv. Skin Wound Care 22, 68–71. doi: 10.1097/01.ASW.0000345283.05532.9a
Soffer, A. (1976). Chihuahua e laetrile, terapia chelante e miele da Boulder, Colorado [editorial]. Arch. Intern. Med.136, 865–866. doi: 10.1001/archinte.136.8.865
Sojka, M., Valachova, I., Bucekova, M., e Majtan, J. (2016). Efficacia antibiofilm del miele e della defensina-1 derivata dalle api su biofilm di ferite multispecie. J. Med. Microbiol. doi: 10.1099/jmm.0.000227 [Epub ahead of print].
Stephens, J. M., Schlothauer, R. C., Morris, B. D., Yang, D., Fearnley, L., Greenwood, D. R., et al. (2010). Composti fenolici e metilgliossale in alcuni mieli di Manuka e Kanuka della Nuova Zelanda. Food Chem. 120, 78–86. doi: 10.1016/j.foodchem.2009.09.074
Tan, H. T., Rahman, R. A., Gan, S. H., Halim, A. S., Hassan, S. A., Sulaiman, S. A., et al. (2009). Le proprietà antibatteriche del miele tualang malese contro microrganismi di ferite e enterici in confronto al miele di Manuka. BMC Complement Altern. Med. 9:34. doi: 10.1186/1472-6882-9-34
Watanabe, K., Rahmasari, R., Matsunaga, A., Haruyama, T., e Kobayashi, N. (2014). Effetti antivirali anti-influenzali del miele in vitro: potente alta attività del miele di manuka. Arch. Med. Res. 45, 359–365. doi: 10.1016/j.arcmed.2014.05.006
Wilkinson, J. M., e Cavanagh, H. M. A. (2005). Attività antibatterica di 13 mieli contro Escherichia coli e Pseudomonas aeruginosa. J. Med. Food 8, 100–103. doi: 10.1089/jmf.2005.8.100
Williams, S., King, J., Revell, M., Manley-Harris, M., Balks, M., Janusch, F., et al. (2014). Variazioni regionali, annuali e individuali nel contenuto di diidrossiacetone nel nettare di manuka (Leptospermum scoparium) in Nuova Zelanda. J. Agric. Food Chem. 62, 10332–10340. doi: 10.1021/jf5045958
Willix, D. J., Molan, P. C., e Harfoot, C. G. (1992). Un confronto della sensibilità delle specie batteriche che infettano le ferite all’attività antibatterica del miele di manuka e di altri mieli. J. Appl. Bacteriol. 73, 388–394. doi: 10.1111/j.1365-2672.1992.tb04993.x
Windsor, S., Pappalardo, M., Brooks, P., Williams, S., e Manley-Harris, M. (2012). Una nuova analisi conveniente di diidrossiacetone e metilgliossale applicata ai mieli australiani di Leptospermum. J. Pharmacogn. Phytother. 4, 6–11.
Miele di Manuka Terapeutico: Non Più Così Alternativo - https://www.frontiersin.org/articles/10.3389/fmicb.2016.00569/full
Lascia un commento