Miel de Manuka thérapeutique : plus si alternatif

La recherche sur le miel médicinal connaît une renaissance importante. D’un remède folklorique largement rejeté par la médecine conventionnelle comme « alternatif », on observe désormais un intérêt croissant de la part des scientifiques, des praticiens cliniques et du grand public pour les usages thérapeutiques du miel. Plusieurs facteurs expliquent cet engouement : d’abord, la montée de la résistance aux antibiotiques chez de nombreux agents pathogènes bactériens a suscité un intérêt pour le développement et l’utilisation de nouveaux antibactériens ; ensuite, un nombre croissant d’études fiables et de rapports de cas ont démontré que certains miels sont très efficaces pour le traitement des plaies ; de plus, le miel thérapeutique se vend à un prix premium, et l’industrie du miel encourage activement des études qui lui permettront d’en tirer profit ; enfin, la nature très complexe et plutôt imprévisible du miel représente un défi attrayant pour les scientifiques en laboratoire. Dans cet article, nous passons en revue la recherche sur le miel de Manuka, depuis les études observationnelles sur ses effets antimicrobiens jusqu’aux travaux expérimentaux et mécanistiques actuels visant à intégrer le miel dans la médecine conventionnelle. Nous soulignons les lacunes et controverses actuelles dans notre compréhension des mécanismes d’action du miel, et suggérons de nouvelles études qui pourraient faire du miel une alternative qui ne soit plus « alternative ».
Introduction
Le miel a été utilisé comme médicament tout au long de l'histoire de l'humanité. L'un des usages thérapeutiques les plus courants et persistants du miel a été comme pansement pour les plaies, presque certainement en raison de ses propriétés antimicrobiennes. Avec l'avènement des antibiotiques très actifs dans les années 1960, le miel a été considéré comme une « substance sans valeur mais inoffensive » (Soffer, 1976). Cependant, la crise actuelle et croissante de la résistance aux antibiotiques a ravivé l'intérêt pour l'utilisation du miel, à la fois comme agent efficace en soi et comme piste thérapeutique pour développer de nouvelles méthodes de traitement. Le miel est généralement issu du nectar des fleurs et produit par les abeilles, le plus souvent l'abeille européenne Apis mellifera, et constitue un mélange complexe de sucres, d'acides aminés, de composés phénoliques et d'autres substances. Les types de miel issus de différentes plantes à fleurs varient considérablement dans leur capacité à tuer les bactéries, ce qui a compliqué la littérature sur le miel et rendu parfois difficile la reproduction des résultats entre différentes études (Allen et al., 1991 ; Irish et al., 2011). La majorité des études récentes portant sur le mécanisme d'action du miel se sont concentrées sur le miel de Manuka actif bien caractérisé et standardisé, produit par certaines espèces de Leptospermum originaires de Nouvelle-Zélande et d'Australie, qui a été enregistré comme produit de soin des plaies auprès des autorités médicales compétentes. Ainsi, sauf indication contraire, cette revue se concentrera sur le miel de Manuka.
Analyses chimiques du miel de Manuka actif
Le professeur Peter Molan de l'université de Waikato, en Nouvelle-Zélande, a été le premier à signaler l'activité inhabituelle du miel de Manuka et a commencé à tester son action contre un large éventail d'espèces bactériennes au milieu des années 1980. Cependant, bien qu'il soit clair que même de faibles concentrations de miel de Manuka tuent les agents pathogènes bactériens, l'ingrédient actif spécifique responsable de cela est resté insaisissable pendant de nombreuses années. La forte teneur en sucre et le pH bas rendent le miel inhibiteur de la croissance microbienne, mais l'activité persiste lorsque ces facteurs sont dilués à des niveaux négligeables. De nombreux types de miel produisent également du peroxyde d'hydrogène lorsque la glucose oxydase, dérivée de l'abeille, réagit avec le glucose et l'eau. Cependant, dans le miel de Manuka, la production de peroxyde d'hydrogène est relativement faible et peut être neutralisée par la catalase, mais l'activité persiste néanmoins. La cause de cette activité résiduelle, appelée « activité non peroxydase » ou NPA, a finalement été révélée en 2008, lorsque deux laboratoires ont identifié indépendamment le méthylglyoxal (MGO) dans le miel de Manuka (Adams et al., 2008 ; Mavric et al., 2008). Le MGO résulte de la déshydratation spontanée de son précurseur dihydroxyacétone (DHA), un phytochemical naturel présent dans le nectar des fleurs de Leptospermum scoparium, Leptospermum polygalifolium et de certaines espèces apparentées de Leptospermum originaires de Nouvelle-Zélande et d'Australie (Adams et al., 2009 ; Williams et al., 2014 ; Norton et al., 2015). Le MGO peut réagir de manière relativement non spécifique avec des macromolécules telles que l'ADN, l'ARN et les protéines (Adams et al., 2008 ; Mavric et al., 2008 ; Majtan et al., 2014b), et pourrait théoriquement être toxique pour les cellules mammifères (Kalapos, 2008). Cependant, aucune preuve de dommage aux cellules hôtes n'a été observée lorsque le miel de Manuka est consommé par voie orale ou utilisé comme pansement ; en effet, le miel semble stimuler la cicatrisation et réduire les cicatrices lorsqu'il est appliqué sur des plaies (Biglari et al., 2013 ; Majtan, 2014 ; Dart et al., 2015). La manière dont il exerce cette toxicité apparemment sélective envers les cellules bactériennes reste inconnue.
Des niveaux élevés de MGO ou de peroxyde d'hydrogène produisent généralement le miel le plus actif, cependant, la corrélation n'est pas toujours parfaite, ce qui suggère que d'autres composants du miel peuvent moduler son activité (Molan, 2008 ; Kwakman et al., 2011 ; Chen et al., 2012 ; Lu et al., 2013). La défensine-1 de l'abeille, un peptide antimicrobien dérivé des abeilles, est responsable de l'activité dans le miel Revamil, un miel actif produit à partir d'une source non divulguée, mais il semble être modifié structurellement et inactif dans le miel de Manuka (Kwakman et al., 2011 ; Majtan et al., 2012). Le niveau de leptosine, un glycoside trouvé exclusivement dans le miel de Leptospermum, est corrélé à la puissance et peut moduler l'activité antimicrobienne du miel de Manuka (Kato et al., 2012). De même, divers composés phénoliques avec un potentiel antimicrobien peuvent être présents, particulièrement dans les miels de couleur plus foncée, et bien que ces composés se trouvent à des niveaux peu susceptibles d'être inhibiteurs seuls, ils peuvent agir en synergie entre eux ou avec d'autres composants du miel pour produire ou modifier l'activité (Estevinho et al., 2008 ; Stephens et al., 2010). Les phénoliques peuvent également agir comme antioxydants et pourraient être responsables des propriétés anti-inflammatoires et cicatrisantes du miel (Stephens et al., 2010). Il convient de noter que toutes les espèces de Leptospermum ne produisent pas de miel actif, et même au sein des miels de L. scoparium et L. polygalifolium, les niveaux de MGO peuvent varier d'environ 100 à plus de 1200 ppm (Windsor et al., 2012). Une enquête sur l'activité des miels australiens a révélé que le miel provenant de plantes de Leptospermum poussant autour de la frontière entre la Nouvelle-Galles du Sud et le Queensland était particulièrement actif, mais il n'est pas connu si cela est dû à la plante, au sol, au climat ou à d'autres facteurs (Irish et al., 2011).
L'inhibition des pathogènes par le miel
Le miel a été testé in vitro sur une large gamme de pathogènes, en particulier ceux pouvant coloniser la peau, les plaies et les muqueuses, où un traitement topique au miel est possible. À ce jour, les tests in vitro ont montré que le miel de Manuka peut inhiber efficacement tous les pathogènes bactériens problématiques testés (résumé dans le Tableau 1). Il est particulièrement intéressant de noter que les isolats cliniques présentant des phénotypes de résistance multiple aux médicaments (MDR) ne montrent aucune réduction de leur sensibilité au miel, ce qui indique un large spectre d'action unique parmi les antimicrobiens connus (Willix et al., 1992 ; Blair et Carter, 2005 ; George et Cutting, 2007 ; Tan et al., 2009). De plus, les tentatives de génération de souches résistantes au miel en laboratoire n'ont pas abouti et aucun cas d'isolat clinique ayant acquis une résistance au miel n'a été rapporté (Blair et al., 2009 ; Cooper et al., 2010).
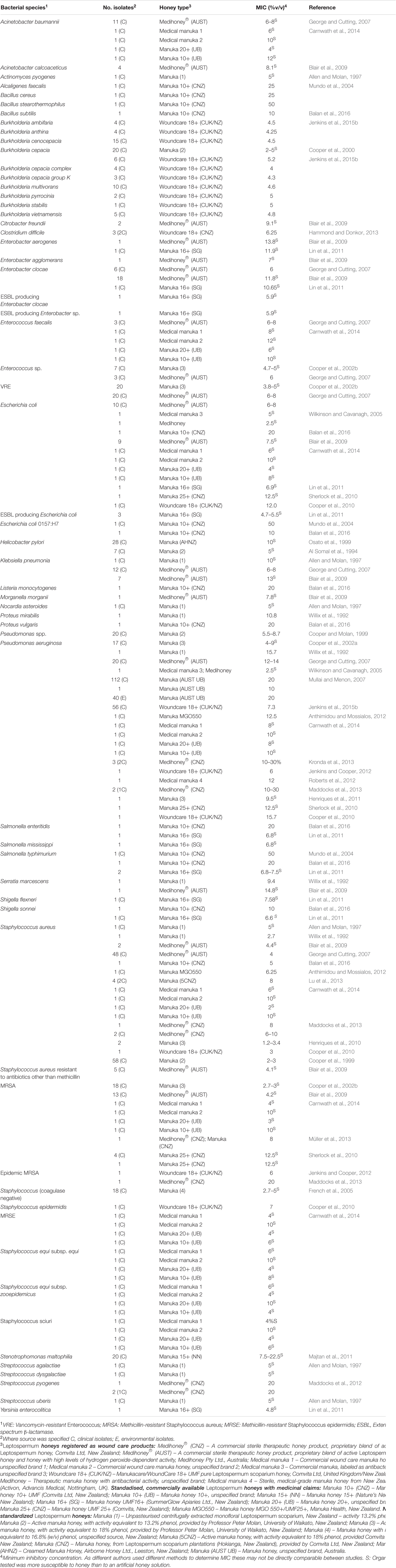
TABLEAU 1. Espèces bactériennes sensibles au miel de Manuka thérapeutique.
En plus d'inhiber les cellules planctoniques, le miel peut disperser et tuer les bactéries vivant dans les biofilms. Les biofilms sont des communautés de cellules généralement enfermées dans une matrice extracellulaire auto-produite et se trouvant adhérées à des surfaces, y compris les plaies, les dents, les surfaces muqueuses et les dispositifs implantés. Les microbes résidant dans les biofilms sont protégés des agents antimicrobiens et peuvent provoquer des infections persistantes et non résolutives. Le miel de Manuka perturbe les agrégats cellulaires (Maddocks et al., 2012 ; Roberts et al., 2012) et empêche la formation de biofilms par un large éventail de pathogènes problématiques, notamment les espèces de Streptococcus et Staphylococcus, Pseudomonas aeruginosa, Escherichia coli, Proteus mirabilis, Enterobacter cloacae, Acinetobacter baumannii et Klebsiella pneumonia (Maddocks et al., 2012, 2013 ; Lu et al., 2014 ; Majtan et al., 2014a ; Halstead et al., 2016). Il est important de noter que le miel peut également perturber les biofilms établis et tuer les cellules résidentes, bien qu'une concentration plus élevée soit nécessaire que pour les cellules planctoniques (Okhiria et al., 2009 ; Maddocks et al., 2013 ; Lu et al., 2014 ; Majtan et al., 2014a). Très récemment, le miel de Manuka a été testé sur un biofilm multispecies contenant Staphylococcus aureus, Streptococcus agalactiae, Pseudomonas aeruginosa et Enterococcus faecalis et il a été constaté qu'il réduisait la viabilité de toutes les espèces sauf E. faecalis, qui n'a pas pu être éradiqué (Sojka et al., 2016). Cela a des implications cliniques évidentes pour l'utilisation du miel sur les plaies contenant des biofilms, et comprendre comment le biofilm permet à E. faecalis de survivre alors qu'il est normalement tué par le miel est un domaine important et intéressant pour de futures études. Le MGO semble être en grande partie, mais pas entièrement, responsable de l'inhibition des biofilms par le miel de Manuka, soulignant à nouveau l'importance des composants supplémentaires qui modulent l'activité (Kilty et al., 2011 ; Lu et al., 2014).
Le spectre d'activité du miel contre les agents pathogènes non bactériens reste à bien définir. Des études récentes examinant l'effet antiviral du miel de Manuka ont suggéré son potentiel pour le traitement du virus varicelle-zona (responsable de la varicelle et du zona) (Shahzad et Cohrs, 2012) et de la grippe (Watanabe et al., 2014). Les agents pathogènes fongiques de la peau, notamment Candida albicans et les espèces dermatophytes, sont nettement moins sensibles que les bactéries au miel de Manuka, mais sont inhibés par le miel produisant de hauts niveaux de peroxyde d'hydrogène (Brady et al., 1996 ; Irish et al., 2006). Le miel de Manuka et non-Manuka a été trouvé pour réduire la viabilité des spores du microsporidien Nosema apis, un pathogène important des abeilles, mais le miel ne pouvait pas guérir l'infection des abeilles une fois celle-ci installée (Malone et al., 2001). Il y a eu très peu d'études sur l'utilisation du miel contre les parasites protozoaires ou helminthes, et celles-ci n'ont pas utilisé de miel avec une activité bien caractérisée, ce qui rend difficile l'évaluation de la portée de leurs résultats (Bassam et al., 1997 ; Nilforoushzadeh et al., 2007 ; Sajid et Azim, 2012).
Intégrer le miel dans la médecine conventionnelle : des études expérimentales et mécanistiques récentes éclairent le mode d'action du miel
Le miel de Manuka actif est largement disponible en tant qu'agent thérapeutique et aliment fonctionnel, et la plupart des consommateurs l'acceptent comme un produit holistique, quelque peu mystérieux. Cependant, un manque de compréhension sur la manière dont le miel tue les bactéries et favorise la guérison limite son acceptation par la médecine conventionnelle, où il est encore considéré comme « alternatif » ou « complémentaire ». La grande majorité des études sur le miel à ce jour ont été descriptives, mais des recherches récentes tentent de démêler le mode d'action du miel en utilisant des approches mécanistiques pour déterminer comment il agit au niveau cellulaire et moléculaire.
Études ultrastructurales des cellules et communautés bactériennes traitées par le miel
Le miel peut profondément modifier la taille et la forme des cellules bactériennes, bien que l'ampleur de ces modifications varie selon les espèces bactériennes. En utilisant la microscopie électronique en transmission (MET), les cultures de S. aureus traitées avec du miel de Manuka présentaient plus de cellules avec des septa complétés comparées à celles traitées avec du miel artificiel, ce qui suggère que les cellules entraient dans la phase de division mais ne parvenaient pas à la terminer, bien que ces cellules paraissent normales en apparence sous microscopie électronique à balayage (MEB) (Henriques et al., 2010). Plus récemment, l'imagerie en contraste de phase après traitement avec une dose sublétale de miel de Manuka a montré que les cellules de S. aureus et Bacillus subtilis étaient significativement plus petites et avaient plus souvent de l'ADN condensé que celles cultivées sans miel (Lu et al., 2013). Il est difficile de comparer directement ces études car elles ont utilisé des quantités de miel et des temps de traitement différents, mais globalement les résultats suggèrent un découplage de la croissance et de la division cellulaire, phénomène souvent observé en réponse à des stress nutritionnels et environnementaux (Silva-Rocha et de Lorenzo, 2010).
Il a été rapporté que le traitement au miel provoque dans les cultures des espèces Gram négatives E. coli et P. aeruginosa des cellules anormalement plus courtes et plus longues (Lu et al., 2013). Fait intéressant, bien que P. aeruginosa semble moins sensible à l'inhibition par le miel que d'autres espèces, des changements cellulaires profonds ont été observés par MET et MEB, incluant des sillons et des boursouflures (protrusions des membranes plasmatiques cellulaires) à la surface cellulaire ainsi qu'une quantité importante de débris extracellulaires indiquant une lyse cellulaire (Henriques et al., 2011). Cela a été confirmé dans une étude ultérieure utilisant la coloration fluorescente BacLight vivant-mort et la microscopie confocale, bien que cette étude ait également montré qu'un nombre relativement important de cellules vivantes subsistait. Ces études ont utilisé du miel à 20 % (p/v), ce qui était supérieur à la CMB pour leur souche de P. aeruginosa et une inhibition et une mortalité substantielles étaient donc attendues. Cependant, la microscopie à force atomique (AFM) utilisant des concentrations sub-bactéricide a tout de même révélé une déformation cellulaire importante et des boursouflures dans les cellules traitées avec des concentrations MIC (12 %) et demi-MIC (6 %), ainsi qu'une lyse cellulaire importante (Roberts et al., 2012). Cette dégénérescence apparente de la cellule de P. aeruginosa a été corroborée par une analyse par PCR quantitative montrant une régulation à la baisse par 10 fois dans les cellules traitées au miel du gène oprF, qui code pour une porine de la membrane externe importante pour la stabilité structurelle (Jenkins et al., 2015a).
Les analyses « omiques » évaluent la réponse globale de la cellule à l'inhibition par le miel
La capacité à évaluer les productions cellulaires globales a révolutionné l’étude des interactions médicament-pathogène et présente une valeur particulière pour des produits naturels complexes comme le miel, où les effets sur plusieurs processus sont probables. Des études par microarrays et protéomiques sur des bactéries exposées au miel ont suggéré une induction des processus liés au stress et une suppression de la synthèse protéique (Blair et al., 2009; Jenkins et al., 2011; Packer et al., 2012). Bien que globalement cela soit assez typique d’une réponse à des agents inhibiteurs, le miel a produit une « signature » unique d’expression différentielle incluant de nombreuses protéines aux fonctions hypothétiques ou inconnues, suggérant un mode d’action nouveau. Des gènes ou protéines spécifiques trouvés comme régulés à la baisse dans les analyses « omiques » de S. aureus et E. coli O157/H7 ont des fonctions liées à la virulence, au quorum sensing et à la formation de biofilm (Lee et al., 2011; Jenkins et al., 2013), et chez P. aeruginosa il y avait une régulation à la baisse des protéines impliquées dans la flagellation (Roberts et al., 2015). Ces phénotypes sont essentiels pour que les pathogènes s’établissent et produisent une infection invasive, ce qui indique qu’en plus d’inhiber la croissance, le miel peut réduire le potentiel pathogène des bactéries infectantes.
Bien que encore relativement limitées en nombre et en portée, les analyses « omiques » réalisées à ce jour suggèrent une réponse cellulaire complexe au miel avec une variation considérable selon les espèces bactériennes. Des approches avancées de biologie des systèmes permettant de contextualiser les données, ainsi que des études de validation utilisant la PCR quantitative et des souches à gènes supprimés, sont désormais nécessaires pour démêler cette complexité, ce qui pourrait révéler de nouvelles approches pour des thérapies médicamenteuses visant à inhiber la croissance bactérienne (Hudson et al., 2012).
Interactions entre le miel et les antibiotiques conventionnels
En plus d'être utilisé comme agent unique, le miel peut également être employé pour augmenter le traitement avec des antibiotiques conventionnels. Cela peut être particulièrement utile lorsqu'il est combiné avec des agents systémiques pouvant être délivrés à un lit de plaie via la circulation sanguine, tandis que le miel est appliqué localement. Les traitements combinés peuvent aussi réduire la dose thérapeutique des agents antimicrobiens et prévenir le développement de résistances, et dans certains cas, peuvent entraîner une synergie médicamenteuse, où l'activité combinée est supérieure à la somme des activités individuelles de chaque partenaire médicamenteux.
Des études in vitro combinant du miel de Manuka thérapeutiquement approuvé avec des agents antibiotiques ont trouvé un effet synergique avec l'oxacilline, la tétracycline, l'imipénem et la mupirocine contre la croissance d'une souche MRSA (Jenkins et Cooper, 2012). De plus, la présence d'une concentration sous-inhibitrice de miel en combinaison avec l'oxacilline a restauré la sensibilité de la souche MRSA à l'oxacilline. Les auteurs ont observé une régulation à la baisse de mecR1, qui code pour une protéine de liaison à la pénicilline spécifique au MRSA (PBP2A), et ont suggéré ce mécanisme comme base de la synergie du miel. Une forte activité synergique entre le miel de Manuka et la rifampicine contre plusieurs souches de S. aureus, y compris des isolats cliniques et des souches MRSA, a également été démontrée, et la présence de miel a empêché l'apparition de résistance à la rifampicine in vitro (Müller et al., 2013). Cela est cliniquement significatif car la rifampicine pénètre bien dans les tissus et les abcès et est couramment utilisée pour traiter les infections staphylococciques superficielles, mais induit rapidement une résistance et doit donc être utilisée en combinaison avec un autre agent. Une autre conclusion de cette étude est que la synergie n'était pas due au MGO, car un miel synthétique enrichi en MGO n'était pas synergique avec la rifampicine.
Comprendre comment le miel affecte l'action des antimicrobiens aux modes d'action bien caractérisés pourrait également approfondir notre compréhension de l'effet du miel sur les agents pathogènes bactériens. Liu et al. (2014) ont étendu l'analyse de la synergie pour inclure des antibiotiques supplémentaires et différentes souches de S. aureus et MRSA. Ils ont suggéré qu'une susceptibilité accrue à la clindamycine et à la gentamicine pourrait résulter de l'effet combiné d'une synthèse protéique régulée à la baisse par le miel avec l'inhibition des ribosomes par les antibiotiques, tandis que la synergie avec les antibiotiques β-lactamines pourrait être due à un stress oxydatif accru causé par les deux partenaires. Comme les souches de S. aureus et MRSA étaient également sensibles à la combinaison oxacilline-miel, il semblait que la synergie n'était probablement pas due à une régulation à la baisse de PBP2A. Cependant, dans un isolat clinique de MRSA, il n'y avait pas d'augmentation de la sensibilité à la clindamycine ou à la gentamicine en présence de miel, ce qui est notable car c'est le premier cas rapporté d'une différence de réponse au miel entre MRSA et S. aureus. Étudier cette différence spécifique à la souche par des analyses transcriptomiques ou protéomiques serait une piste intéressante pour de futures recherches (Liu et al., 2014).
Preuves d'efficacité issues d'études animales, de rapports de cas et d'essais cliniques
Les entreprises qui produisent et commercialisent le miel de Manuka promeuvent des normes éthiques élevées et découragent l'utilisation de modèles animaux pour étudier les infections et la cicatrisation des plaies. Le miel de Manuka a cependant été utilisé pour traiter des animaux avec des plaies chirurgicales ou accidentelles, en particulier des chevaux, avec des résultats positifs (Dart et al., 2015 ; Bischofberger et al., 2016). Des rapports de cas utilisant le miel pour des plaies et ulcères non cicatrisants ont noté une amélioration significative avec résolution de l'infection là où les antibiotiques conventionnels avaient échoué (Regulski, 2008 ; Smith et al., 2009). Cependant, malgré cela et les preuves issues de nombreux modèles in vitro et in vivo montrant que le miel tue les pathogènes problématiques des plaies, il existe une rareté de données cliniques solides pour le miel de Manuka. Plusieurs raisons expliquent cela, notamment les difficultés techniques à réaliser un essai contrôlé en double aveugle avec placebo sur une substance distinctive comme le miel, des considérations éthiques, le manque d'intérêt des praticiens cliniques et le rapport coût-bénéfice pour les entreprises de miel, dont l'objectif est les produits naturels et les ventes en libre-service où le miel de Manuka et les pansements associés ont déjà un prix premium. Ces facteurs pourraient évoluer à mesure que la résistance aux antibiotiques réduit les options de traitement actuelles et que la recherche continue mettant en lumière le potentiel du miel attire l'attention des praticiens médicaux.
Lacunes et opportunités émergentes dans l'étude du miel
De grands progrès ont été réalisés récemment dans notre compréhension du miel thérapeutique, pourtant son utilisation en médecine clinique reste limitée, même lorsque les antibiotiques conventionnels commencent à échouer. La complexité du miel, qui est sans doute sa plus grande force pour tuer divers pathogènes et prévenir la résistance, complique son étude car de nombreux facteurs agissant ensemble influencent probablement son activité. Nous préconisons des études mécanistiques supplémentaires utilisant du miel de Manuka thérapeutique dûment enregistré, en particulier des études employant des approches de biologie systémique non réductionnistes, ainsi que des analyses chimiques et microbiologiques détaillées pour élucider comment le miel agit au niveau moléculaire, cellulaire et de la population, comment cela peut différer selon les souches et espèces de pathogènes microbiens, et comment la cellule hôte réagit (Tableau 2). Les informations obtenues de ces études peuvent ensuite orienter la thérapie et produire les données cliniques nécessaires pour intégrer le miel dans la médecine conventionnelle ; il ne s'agit plus d'une thérapie alternative utilisée uniquement en dernier recours.
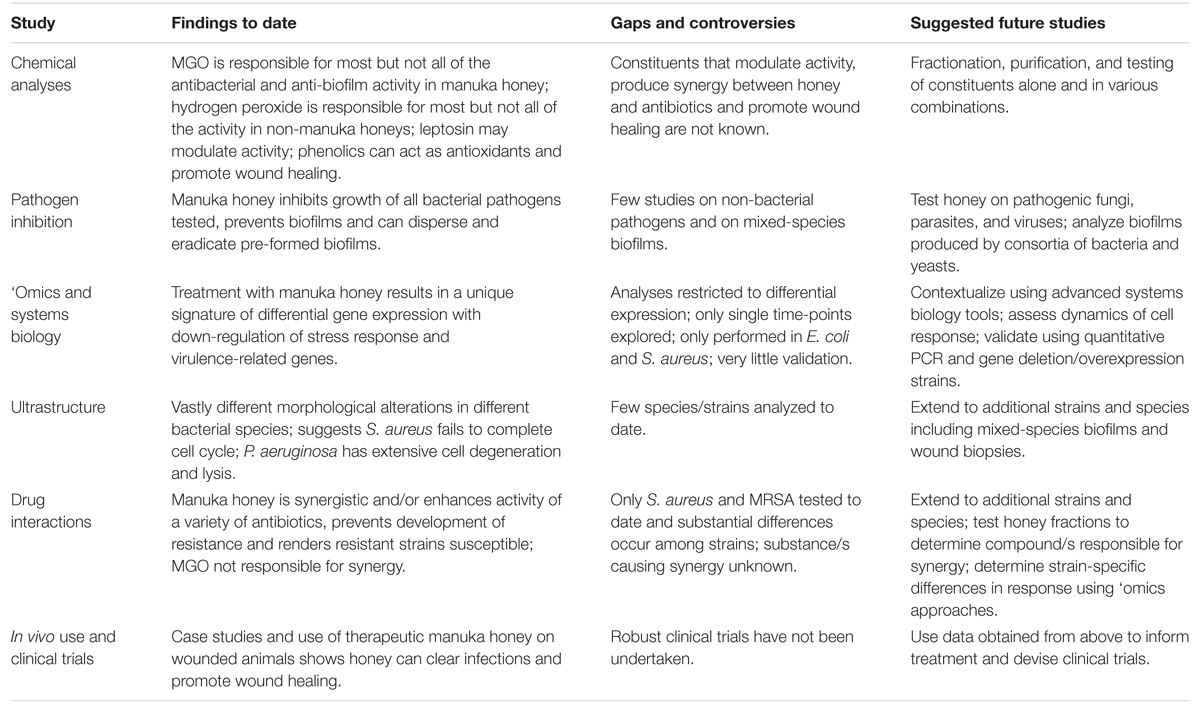
TABLEAU 2. Études sur le miel de Manuka : résultats, lacunes et études futures.
Contributions des auteurs
Cette revue a été rédigée par DC, SB, NNC, DB et PB et a été critiquement revue par RS et EH.
Financement
NNC reçoit un soutien salarial de la Rural Industries Research and Development Corporation – Honey Bee Program (subvention PRJ-009186).
Déclaration de conflit d'intérêts
DC, PB, et EH déclarent un soutien financier et non financier sous forme de miel de Manuka de Comvita NZ Limited et Capilano Honey Limited ; RS est employé par Comvita NZ Limited, qui commercialise du miel de Manuka de qualité médicale (Medihoney).
Les autres auteurs déclarent que la recherche a été conduite en l'absence de toute relation commerciale ou financière pouvant être interprétée comme un conflit d'intérêts potentiel.
Abréviation
ESBL, β-lactamase à spectre étendu ; MBC, concentration bactéricide minimale ; MGO, methyl glyoxal ; MIC, concentration inhibitrice minimale ; MRSA, Staphylococcus aureus résistant à la méthicilline ; MRSE, Staphylococcus epidermis résistant à la méthicilline ; NPA, activité non peroxydase ; VRE, Enterococcus résistant à la vancomycine.
Références
Adams, C. J., Boult, C. H., Deadman, B. J., Farr, J. M., Grainger, M. N. C., Manley-Harris, M., et al. (2008). Isolement par HPLC et caractérisation de la fraction bioactive du miel de Manuka (Leptospermum scoparium) de Nouvelle-Zélande. Carbohydr. Res. 343, 651–659. doi: 10.1016/j.carres.2007.12.011
Adams, C. J., Manley-Harris, M., et Molan, P. C. (2009). L'origine du methylglyoxal dans le miel de Manuka (Leptospermum scoparium) de Nouvelle-Zélande. Carbohydr. Res. 344, 1050–1053. doi: 10.1016/j.carres.2009.03.020
Al Somal, N., Coley, K. E., Molan, P. C., et Hancock, B. M. (1994). Sensibilité de Helicobacter pylori à l'activité antibactérienne du miel de Manuka. J. R. Soc. Med. 87, 9–12.
Allen, K., Molan, P., et Reid, G. (1991). Une enquête sur l'activité antibactérienne de certains miels de Nouvelle-Zélande. J. Pharm. Pharmacol.43, 817–822. doi: 10.1111/j.2042-7158.1991.tb03186.x
Allen, K. L., et Molan, P. C. (1997). Sensibilité des bactéries responsables de la mammite à l'activité antibactérienne du miel. N. Z. J. Agric. Res. 40, 537–540. doi: 10.1080/00288233.1997.9513276
Anthimidou, E., et Mossialos, D. (2012). Activité antibactérienne des miels grecs et chypriotes contre Staphylococcus aureus et Pseudomonas aeruginosa en comparaison avec le Manuka honey. J. Med. Food 16, 42–47. doi: 10.1089/jmf.2012.0042
Balan, P., Mal, G., Das, S., et Singh, H. (2016). Activités antimicrobiennes synergiques et additives de la curcumine, du Manuka honey et des protéines de lactosérum. J. Food Biochem. doi: 10.1111/jfbc.12249
Bassam, Z., Zohra, B. I., et Saada, A.-A. (1997). Effets du miel sur les parasites Leishmania : une étude in vitro. Trop. Doctor27, 36–38.
Biglari, B., Moghaddam, A., Santos, K., Blaser, G., Büchler, A., Jansen, G., et al. (2013). Étude observationnelle prospective multicentrique sur les soins professionnels des plaies utilisant du miel (Medihoney). Int. Wound J. 10, 252–259. doi: 10.1111/j.1742-481X.2012.00970.x
Bischofberger, A., Dart, C., Horadagoda, N., Perkins, N., Jeffcott, L., Little, C., et al. (2016). Effet du gel de Manuka honey sur les concentrations de facteur de croissance transformant β1 et β3, le nombre de bactéries et l'histomorphologie des plaies cutanées contaminées en pleine épaisseur sur les membres distaux équins. Aust. Vet. J. 94, 27–34. doi: 10.1111/avj.12405
Blair, S., Cokcetin, N., Harry, E., et Carter, D. (2009). L'activité antibactérienne inhabituelle du miel médical de Leptospermum : spectre antibactérien, résistance et analyse du transcriptome. Eur. J. Clin. Microbiol. Infect. Dis. 28, 1199–1208. doi: 10.1007/s10096-009-0763-z
Blair, S. E., et Carter, D. A. (2005). Le potentiel du miel dans la gestion des plaies et des infections. J. Austral. Infect. Control 10, 24–31.
Brady, N., Molan, P., et Harfoot, C. (1996). La sensibilité des dermatophytes à l'activité antimicrobienne du miel de Manuka et d'autres miels. Pharm. Pharmacol. Commun. 2, 471–473.
Carnwath, R., Graham, E. M., Reynolds, K., et Pollock, P. J. (2014). L'activité antimicrobienne du miel contre des isolats bactériens courants de plaies équines. Vet. J. 199, 110–114. doi: 10.1016/j.tvjl.2013.07.003
Chen, C., Campbell, L., Blair, S. E., et Carter, D. A. (2012). L'effet du traitement thermique sur les propriétés antimicrobiennes du miel. Front. Microbiol. 3:265. doi: 10.3389/fmicb.2012.00265
Cooper, R., Jenkins, L., Henriques, A., Duggan, R., et Burton, N. (2010). Absence de résistance bactérienne au miel médical de Manuka. Eur. J. Clin. Microbiol. Infect. Dis. 29, 1237–1241. doi: 10.1007/s10096-010-0992-1
Cooper, R. A., Halas, E., et Molan, P. C. (2002a). L'efficacité du miel pour inhiber des souches de Pseudomonas aeruginosa provenant de brûlures infectées. J. Burn Care Rehabil. 23, 366–370. doi: 10.1097/00004630-200211000-00002
Cooper, R. A., et Molan, P. C. (1999). Utilisation du miel comme antiseptique dans la gestion des infections à Pseudomonas. J. Wound Care 8, 161–164. doi: 10.12968/jowc.1999.8.4.25867
Cooper, R. A., Molan, P. C., et Harding, K. G. (1999). Activité antibactérienne du miel contre des souches de Staphylococcus aureus issues de plaies infectées. J. R. Soc. Med. 92, 283–285.
Cooper, R. A., Molan, P. C., et Harding, K. G. (2002b). Sensibilité au miel des cocci Gram-positifs d'importance clinique isolés de plaies. J. Appl. Microbiol. 93, 857–863. doi: 10.1046/j.1365-2672.2002.01761.x
Cooper, R. A., Wigley, P., et Burton, N. F. (2000). Sensibilité des souches multirésistantes de Burkholderia cepacia au miel. Lett. Appl. Microbiol. 31, 20–24. doi: 10.1046/j.1472-765x.2000.00756.x
Dart, A., Bischofberger, A., Dart, C., et Jeffcott, L. (2015). Revue des recherches sur la cicatrisation des plaies équines par seconde intention utilisant le miel de Manuka : recommandations actuelles et applications futures. Equine Vet. Educ. 27, 658–664. doi: 10.1111/eve.12379
Estevinho, L., Pereira, A. P., Moreira, L., Dias, L. G., et Pereira, E. (2008). Effets antioxydants et antimicrobiens des extraits de composés phénoliques du miel du Nord-Est du Portugal. Food Chem. Toxicol. 46, 3774–3779. doi: 10.1016/j.fct.2008.09.062
French, V. M., Cooper, R. A., et Molan, P. C. (2005). L’activité antibactérienne du miel contre les Staphylocoques à coagulase négative. J. Antimicrobial Chemother. 56, 228–231. doi : 10.1093/jac/dki193
George, N. M., et Cutting, K. F. (2007). Miel antibactérien (Medihoney) : activité in vitro contre des isolats cliniques de MRSA, VRE et autres organismes gram-négatifs multirésistants, y compris Pseudomonas aeruginosa. Wounds 19:231.
Halstead, F. D., Webber, M. A., Rauf, M., Burt, R., Dryden, M., et Oppenheim, B. A. (2016). Activité in vitro d’un miel modifié, de miels médicaux et de pansements antimicrobiens contre des isolats bactériens cliniques producteurs de biofilm. J. Wound Care 25, 93–102. doi : 10.12968/jowc.2016.25.2.93
Hammond, E. N., et Donkor, E. S. (2013). Effet antibactérien du miel de Manuka sur Clostridium difficile. BMC Res. 6:188. doi : 10.1186/1756-0500-6-188
Henriques, A. F., Jenkins, R. E., Burton, N. F., et Cooper, R. A. (2010). Les effets intracellulaires du miel de Manuka sur Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 29, 45–50. doi : 10.1007/s10096-009-0817-2
Henriques, A. F., Jenkins, R. E., Burton, N. F., et Cooper, R. A. (2011). L'effet du miel de Manuka sur la structure de Pseudomonas aeruginosa. Eur. J. Clin. Microbiol. Infect. Dis. 30, 167–171. doi : 10.1007/s10096-010-1065-1
Hudson, N. J., Dalrymple, B. P., et Reverter, A. (2012). Au-delà de l'expression différentielle : la quête des mutations causales et des molécules effectrices. BMC Genomics 13:356. doi : 10.1186/1471-2164-13-356
Irish, J., Blair, S., et Carter, D. (2011). L'activité antibactérienne du miel dérivé de la flore australienne. PLoS ONE 6:e18229. doi : 10.1371/journal.pone.0018229
Irish, J., Carter, D. A., Shokohi, T., et Blair, S. E. (2006). Le miel a un effet antifongique contre les espèces de Candida. Med. Mycol. 44, 289–291. doi : 10.1080/13693780600931986
Jenkins, R., Burton, N., et Cooper, R. (2011). Effet du miel de Manuka sur l'expression de la protéine universelle de stress A chez Staphylococcus aureus résistant à la méthicilline. Int. J. Antimicrob. Agents 37, 373–376. doi : 10.1016/j.ijantimicag.2010.11.036
Jenkins, R., Burton, N., et Cooper, R. (2013). Analyse protéomique et génomique de Staphylococcus aureus résistant à la méthicilline (MRSA) exposé au miel de Manuka in vitro démontrant une régulation à la baisse des marqueurs de virulence. J. Antimicrobial Chemother. 69, 603–615. doi : 10.1093/jac/dkt430
Jenkins, R., et Cooper, R. (2012). Amélioration de l’activité antibiotique contre les agents pathogènes des plaies avec le miel de Manuka in vitro. PLoS ONE 7:e45600. doi : 10.1371/journal.pone.0045600
Jenkins, R., Roberts, A., et Brown, H. L. (2015a). Sur les effets antibactériens du miel de Manuka : perspectives mécanistiques. Res. Rep. Biol. 6, 215–224. doi : 10.2147/RRB.S75754
Jenkins, R., Wootton, M., Howe, R., et Cooper, R. (2015b). Une démonstration de la sensibilité des isolats cliniques obtenus de patients atteints de fibrose kystique au miel de Manuka. Arch. Microbiol. 197, 597–601. doi : 10.1007/s00203-015-1091-6
Kalapos, M. P. (2008). Le tandem des radicaux libres et du méthylglyoxal. Chem. Biol. Interact. 171, 251–271. doi : 10.1016/j.cbi.2007.11.009
Kato, Y., Umeda, N., Maeda, A., Matsumoto, D., Kitamoto, N., et Kikuzaki, H. (2012). Identification d’un nouveau glycoside, leptosine, comme marqueur chimique du miel de Manuka. J. Agric. Food Chem. 60, 3418–3423. doi : 10.1021/jf300068w
Kilty, S. J., Duval, M., Chan, F. T., Ferris, W., et Slinger, R. (2011). Méthylglyoxal : (agent actif du miel de Manuka) activité in vitro contre les biofilms bactériens. Int. Forum Allergy Rhinol. 1, 348–350. doi : 10.1002/alr.20073
Kronda, J. M., Cooper, R. A., et Maddocks, S. E. (2013). Le miel de Manuka inhibe la production de sidérophores chez Pseudomonas aeruginosa. J. Appl. Microbiol. 115, 86–90. doi: 10.1111/jam.12222
Kwakman, P. H., te Velde, A. A., de Boer, L., Vandenbroucke-Grauls, C. M., et Zaat, S. A. (2011). Deux miels médicinaux majeurs ont des mécanismes différents d'activité bactéricide. PLoS ONE 6:e17709. doi: 10.1371/journal.pone.0017709
Lee, J.-H., Park, J.-H., Kim, J.-A., Neupane, G. P., Cho, M. H., Lee, C.-S., et al. (2011). De faibles concentrations de miel réduisent la formation de biofilm, la communication cellulaire (quorum sensing) et la virulence chez Escherichia coli O157: H7. Biofouling 27, 1095–1104. doi: 10.1080/08927014.2011.633704
Lin, S. M., Molan, P. C., et Cursons, R. T. (2011). La sensibilité contrôlée in vitro des agents pathogènes gastro-intestinaux à l'effet antibactérien du miel de Manuka. Eur. J. Clin. Microbiol. Infect. Dis. 30, 569–574. doi: 10.1007/s10096-010-1121-x
Liu, M., Lu, J., Müller, P., Turnbull, L., Burke, C. M., Schlothauer, R. C., et al. (2014). Différences spécifiques aux antibiotiques dans la réponse de Staphylococcus aureus au traitement avec des antimicrobiens combinés au miel de Manuka. Front. Microbiol. 5:779. doi: 10.3389/fmicb.2014.00779
Lu, J., Carter, D. A., Turnbull, L., Rosendale, D., Hedderley, D., Stephens, J., et al. (2013). L'effet des miels de kanuka, Manuka et trèfle de Nouvelle-Zélande sur la dynamique de croissance bactérienne et la morphologie cellulaire varie selon l'espèce. PLoS ONE8:e55898. doi : 10.1371/journal.pone.0055898
Lu, J., Turnbull, L., Burke, C. M., Liu, M., Carter, D. A., Schlothauer, R. C., et al. (2014). Les miels de type Manuka peuvent éradiquer les biofilms produits par des souches de Staphylococcus aureus avec différentes capacités de formation de biofilm. PeerJ 2:e326. doi : 10.7717/peerj.326
Maddocks, S. E., Jenkins, R. E., Rowlands, R. S., Purdy, K. J., et Cooper, R. A. (2013). Le miel de Manuka inhibe l'adhésion et l'invasion des bactéries de plaies médicalement importantes in vitro. Fut. Microbiol. 8, 1523–1536. doi : 10.2217/fmb.13.126
Maddocks, S. E., Lopez, M. S., Rowlands, R. S., et Cooper, R. A. (2012). Le miel de Manuka inhibe le développement des biofilms de Streptococcus pyogenes et réduit l'expression de deux protéines de liaison à la fibronectine. Microbiology 158, 781–790. doi : 10.1099/mic.0.053959-0
Majtan, J. (2014). Le miel : un immunomodulateur dans la cicatrisation des plaies. Wound Repair Regenerat. 22, 187–192. doi : 10.1111/wrr.12117
Majtan, J., Bohova, J., Horniackova, M., Klaudiny, J., et Majtan, V. (2014a). Effets anti-biofilm du miel contre les agents pathogènes des plaies Proteus mirabilis et Enterobacter cloacae. Phytother. Res. 28, 69–75. doi : 10.1002/ptr.4957
Majtan, J., Bohova, J., Prochazka, E., et Klaudiny, J. (2014b). Le méthylglyoxal peut affecter l'accumulation de peroxyde d'hydrogène dans le miel de Manuka par l'inhibition de la glucose oxydase. J. Med. Food 17, 290–293. doi : 10.1089/jmf.2012.0201
Majtan, J., Klaudiny, J., Bohova, J., Kohutova, L., Dzurova, M., Sediva, M., et al. (2012). Modifications induites par le méthylglyoxal des composants protéiques importants de l'abeille dans le miel de Manuka : implications thérapeutiques possibles. Fitoterapia 83, 671–677. doi : 10.1016/j.fitote.2012.02.002
Majtan, J., Majtanova, L., Bohova, J., et Majtan, V. (2011). Le miel de miellat comme agent antibactérien puissant dans l'éradication d'isolats multi-résistants de Stenotrophomonas maltophilia chez des patients atteints de cancer. Phytother. Res. 25, 584–587. doi : 10.1002/ptr.3304
Malone, L. A., Gatehouse, H. S., et Tregidga, E. L. (2001). Effets du temps, de la température et du miel sur Nosema apis (Microsporidia : Nosematidae), un parasite de l'abeille domestique, Apis mellifera (Hymenoptera : Apidae). J. Invertebrate Pathol. 77, 258–268. doi : 10.1006/jipa.2001.5028
Mavric, E., Wittmann, S., Barth, G., et Henle, T. (2008). Identification et quantification du méthylglyoxal comme principal constituant antibactérien des miels de Manuka (Leptospermum scoparium) de Nouvelle-Zélande. Mol. Nutrit. Food Res. 52, 483–489. doi: 10.1002/mnfr.200700282
Molan, P. M. (2008). Une explication de pourquoi le niveau de MGO dans le miel de Manuka ne reflète pas l’activité antibactérienne. New Zealand Beekeeper 16, 11–13.
Mullai, V., et Menon, T. (2007). Activité bactéricide de différents types de miel contre des isolats cliniques et environnementaux de Pseudomonas aeruginosa. J. Alternat. Complement. Med. 13, 439–442. doi: 10.1089/acm.2007.6366
Müller, P., Alber, D. G., Turnbull, L., Schlothauer, R. C., Carter, D. A., Whitchurch, C. B., et al. (2013). Synergie entre Medihoney et la rifampicine contre le Staphylococcus aureus résistant à la méthicilline (MRSA). PLoS ONE 8:e57679. doi: 10.1371/journal.pone.0057679
Mundo, M. A., Padilla-Zakour, O. I., et Worobo, R. W. (2004). Inhibition de la croissance des agents pathogènes alimentaires et des organismes de détérioration des aliments par certains miels bruts. Int. J. Food Microbiol. 97, 1–8. doi: 10.1016/j.ijfoodmicro.2004.03.025
Nilforoushzadeh, M. A., Jaffary, F., Moradi, S., Derakhshan, R., et Haftbaradaran, E. (2007). Effet de l’application topique de miel associée à une injection intralésionnelle de glucantime dans le traitement de la leishmaniose cutanée. BMC Complement Altern. Med. 7:1. doi: 10.1186/1472-6882-7-1
Norton, A. M., McKenzie, L. N., Brooks, P. R., et Pappalardo, L. J. (2015). Quantification de la dihydroxyacétone dans le nectar australien de Leptospermum par chromatographie liquide à haute performance. J. Agric. Food Chem. 63, 6513–6517. doi: 10.1021/acs.jafc.5b01930
Okhiria, O., Henriques, A., Burton, N., Peters, A., et Cooper, R. (2009). Le miel module les biofilms de Pseudomonas aeruginosa de manière dépendante du temps et de la dose. J. ApiProduct. ApiMedical Sci. 1, 6–10. doi: 10.3896/IBRA.4.01.1.03
Osato, M. S., Reddy, S. G., et Graham, D. Y. (1999). Effet osmotique du miel sur la croissance et la viabilité de Helicobacter pylori. Dig. Dis. Sci. 44, 462–464. doi: 10.1023/A:1026676517213
Packer, J. M., Irish, J., Herbert, B. R., Hill, C., Padula, M., Blair, S. E., et al. (2012). Effet antibactérien spécifique non peroxydique du miel de Manuka sur le protéome de Staphylococcus aureus. Int. J. Antimicrob. Agents 40, 43–50. doi: 10.1016/j.ijantimicag.2012.03.012
Regulski, M. (2008). Un nouveau pansement pour le traitement des ulcères chroniques des jambes. Podiatry Manag. 27, 235–246.
Roberts, A. E., Maddocks, S. E., et Cooper, R. A. (2012). Le miel de Manuka est bactéricide contre Pseudomonas aeruginosa et induit une expression différentielle de oprF et algD. Microbiology 158, 3005–3013. doi : 10.1099/mic.0.062794-0
Roberts, A. E. L., Maddocks, S. E., et Cooper, R. A. (2015). Le miel de Manuka réduit la motilité de Pseudomonas aeruginosa par suppression des gènes associés aux flagelles. J. Antimicrob. Chemother. 70, 716–725. doi : 10.1093/jac/dku448
Sajid, M., et Azim, M. K. (2012). Caractérisation de l'activité nématicide du miel naturel. J. Agric. Food Chem. 60, 7428–7434. doi : 10.1021/jf301653n
Shahzad, A., et Cohrs, R. J. (2012). Activité antivirale in vitro du miel contre le virus varicelle-zona (VZV) : une étude de médecine translationnelle pour un remède potentiel contre le zona. Transl. Biomed. 3:2.
Sherlock, O., Dolan, A., Athman, R., Power, A., Gethin, G., Cowman, S., et al. (2010). Comparaison de l'activité antimicrobienne du miel d'Ulmo du Chili et du miel de Manuka contre Staphylococcus aureus résistant à la méthicilline, Escherichia coli et Pseudomonas aeruginosa. BMC Complement Altern. Med. 10:47. doi : 10.1186/1472-6882-10-47
Silva-Rocha, R., et de Lorenzo, V. (2010). Bruit et robustesse dans les réseaux de régulation procaryotes. Annu. Rev. Microbiol. 64, 257–275. doi : 10.1146/annurev.micro.091208.073229
Smith, T., Hanft, J. R., et Legel, K. (2009). Miel topique de Leptospermum dans les plaies veineuses récalcitrantes : une série de cas préliminaire. Adv. Skin Wound Care 22, 68–71. doi: 10.1097/01.ASW.0000345283.05532.9a
Soffer, A. (1976). Chihuahuas et laétrile, thérapie de chélation, et miel de Boulder, Colorado [editorial]. Arch. Intern. Med.136, 865–866. doi: 10.1001/archinte.136.8.865
Sojka, M., Valachova, I., Bucekova, M., et Majtan, J. (2016). Efficacité antibiofilm du miel et de la défensine-1 dérivée des abeilles sur un biofilm de plaie multi-espèces. J. Med. Microbiol. doi: 10.1099/jmm.0.000227 [Epub ahead of print].
Stephens, J. M., Schlothauer, R. C., Morris, B. D., Yang, D., Fearnley, L., Greenwood, D. R., et al. (2010). Composés phénoliques et méthylglyoxal dans certains miels de Manuka et Kanuka de Nouvelle-Zélande. Food Chem. 120, 78–86. doi: 10.1016/j.foodchem.2009.09.074
Tan, H. T., Rahman, R. A., Gan, S. H., Halim, A. S., Hassan, S. A., Sulaiman, S. A., et al. (2009). Les propriétés antibactériennes du miel tualang malaisien contre les micro-organismes des plaies et entériques en comparaison avec le miel de Manuka. BMC Complement Altern. Med. 9:34. doi: 10.1186/1472-6882-9-34
Watanabe, K., Rahmasari, R., Matsunaga, A., Haruyama, T., et Kobayashi, N. (2014). Effets antiviraux anti-influenza du miel in vitro : forte activité puissante du miel de manuka. Arch. Med. Res. 45, 359–365. doi : 10.1016/j.arcmed.2014.05.006
Wilkinson, J. M., et Cavanagh, H. M. A. (2005). Activité antibactérienne de 13 miels contre Escherichia coli et Pseudomonas aeruginosa. J. Med. Food 8, 100–103. doi : 10.1089/jmf.2005.8.100
Williams, S., King, J., Revell, M., Manley-Harris, M., Balks, M., Janusch, F., et al. (2014). Variations régionales, annuelles et individuelles de la teneur en dihydroxyacétone du nectar de manuka (Leptospermum scoparium) en Nouvelle-Zélande. J. Agric. Food Chem. 62, 10332–10340. doi : 10.1021/jf5045958
Willix, D. J., Molan, P. C., et Harfoot, C. G. (1992). Comparaison de la sensibilité des espèces bactériennes infectant les plaies à l’activité antibactérienne du miel de manuka et d’autres miels. J. Appl. Bacteriol. 73, 388–394. doi : 10.1111/j.1365-2672.1992.tb04993.x
Windsor, S., Pappalardo, M., Brooks, P., Williams, S., et Manley-Harris, M. (2012). Une nouvelle analyse pratique de la dihydroxyacétone et du méthylglyoxal appliquée aux miels australiens de Leptospermum. J. Pharmacogn. Phytother. 4, 6–11.
Miel de Manuka thérapeutique : plus si alternatif - https://www.frontiersin.org/articles/10.3389/fmicb.2016.00569/full
Laissez un commentaire