Terapeutisk Manuka-honung: Inte längre så alternativ

Forskningen kring medicinskt honung genomgår en betydande renässans. Från att ha varit en folklig huskur som till stor del avfärdats av den etablerade medicinen som "alternativ", ser vi nu ett ökat intresse från forskare, kliniska praktiker och allmänheten för honungens terapeutiska användningsområden. Det finns flera drivkrafter bakom detta intresse: för det första har ökningen av antibiotikaresistens hos många bakteriepatogener väckt intresse för att utveckla och använda nya antibakteriella medel; för det andra har ett ökande antal tillförlitliga studier och fallrapporter visat att vissa honungssorter är mycket effektiva vid sårbehandling; för det tredje har terapeutisk honung ett högt pris, och honungsindustrin främjar aktivt studier som kan göra det möjligt att kapitalisera på detta; och slutligen erbjuder honungens mycket komplexa och ganska oförutsägbara natur en attraktiv utmaning för laboratorieforskare. I denna artikel granskar vi Manuka honung-forskning, från observationsstudier om dess antimikrobiella effekter till aktuell experimentell och mekanistisk forskning som syftar till att föra honung in i den etablerade medicinen. Vi beskriver nuvarande kunskapsluckor och återstående kontroverser kring hur honung verkar, och föreslår nya studier som kan göra honung till ett alternativ som inte längre är "alternativt".
Introduktion
Honung har använts som medicin genom hela människans historia. En av de vanligaste och mest bestående terapeutiska användningarna av honung har varit som sårförband, nästan säkert på grund av dess antimikrobiella egenskaper. Med framväxten av mycket aktiva antibiotika på 1960-talet avfärdades honung som ett "värdelöst men ofarligt ämne" (Soffer, 1976). Den nuvarande och växande krisen med antibiotikaresistens har dock återupplivat intresset för användning av honung, både som ett effektivt medel i sig och som en terapeutisk ledtråd för att utveckla nya behandlingsmetoder. Honung utvinns vanligtvis från nektar från blommor och produceras av bin, oftast det europeiska honungsbiet Apis mellifera, och är en komplex blandning av sockerarter, aminosyror, fenoler och andra ämnen. Honungstyper som kommer från olika blommande växter varierar avsevärt i sin förmåga att döda bakterier, vilket har försvårat litteraturen om honung och ibland gjort det svårt att reproducera resultat mellan olika studier (Allen et al., 1991; Irish et al., 2011). Majoriteten av de senaste studierna som undersöker honungens verkningsmekanism har fokuserat på välkarakteriserad, standardiserad aktiv Manuka-honung som produceras av vissa Leptospermum-arter som är inhemska i Nya Zeeland och Australien, och som har registrerats som en sårvårdsprodukt hos relevanta medicinska tillsynsmyndigheter. Således, om inget annat anges, kommer denna översikt att fokusera på Manuka-honung.
Kemiska analyser av aktiv Manuka-honung
Professor Peter Molan vid Waikato University i Nya Zeeland var den första som rapporterade den ovanliga aktiviteten hos Manuka-honung och började testa dess verkan mot ett brett spektrum av olika bakteriearter i mitten av 1980-talet. Det var dock tydligt att även låga koncentrationer av Manuka-honung dödade bakteriepatogener, men den specifika aktiva ingrediensen som låg bakom detta förblev okänd i många år. Högt sockerinnehåll och lågt pH gör honung hämmande för mikrobiell tillväxt, men aktiviteten kvarstår när dessa späds ut till försumbar nivå. Många olika typer av honung producerar också väteperoxid när glukosoxidas, som härstammar från honungsbiet, reagerar med glukos och vatten. I Manuka-honung är dock produktionen av väteperoxid relativt låg och kan neutraliseras av katalas, men aktiviteten kvarstår ändå. Orsaken till denna kvarvarande aktivitet, kallad ”non-peroxide activity” eller NPA, avslöjades slutligen 2008 när två laboratorier oberoende identifierade metylglyoxal (MGO) i Manuka-honung (Adams et al., 2008; Mavric et al., 2008). MGO bildas genom spontan dehydrering av dess föregångare dihydroxyaceton (DHA), en naturligt förekommande fytokemikalie som finns i nektarn från blommor av Leptospermum scoparium, Leptospermum polygalifolium och några närbesläktade Leptospermum-arter som är inhemska i Nya Zeeland och Australien (Adams et al., 2009; Williams et al., 2014; Norton et al., 2015). MGO kan reagera relativt ospecifikt med makromolekyler som DNA, RNA och proteiner (Adams et al., 2008; Mavric et al., 2008; Majtan et al., 2014b) och kan teoretiskt vara giftigt för däggdjursceller (Kalapos, 2008). Det finns dock inga bevis för skador på värdceller när Manuka-honung antingen konsumeras oralt eller används som sårförband; honung verkar snarare stimulera läkning och minska ärrbildning när den appliceras på sår (Biglari et al., 2013; Majtan, 2014; Dart et al., 2015). Hur den utövar denna till synes selektiva toxicitet mot bakterieceller är fortfarande okänt.
Höga nivåer av MGO eller väteperoxid ger vanligtvis den mest aktiva honungen, men korrelationen är inte alltid perfekt vilket tyder på att andra komponenter i honung kan påverka aktiviteten (Molan, 2008; Kwakman et al., 2011; Chen et al., 2012; Lu et al., 2013). Bee defensin-1, en antimikrobiell peptid från bin, är ansvarig för aktiviteten i Revamil-honung, en aktiv honung producerad från en okänd källa, men denna verkar vara strukturellt modifierad och inaktiv i Manuka-honung (Kwakman et al., 2011; Majtan et al., 2012). Nivån av leptosin, en glykosid som endast finns i Leptospermum-honung, korrelerar med styrkan och kan påverka den antimikrobiella aktiviteten i Manuka-honung (Kato et al., 2012). På liknande sätt kan olika fenoliska föreningar med potentiell antimikrobiell aktivitet finnas, särskilt i mörkare honungssorter, och även om dessa förekommer i nivåer som sannolikt inte är hämmande på egen hand kan de samverka med varandra eller andra komponenter i honung för att skapa eller förändra aktiviteten (Estevinho et al., 2008; Stephens et al., 2010). Fenoler kan också fungera som antioxidanter och kan vara ansvariga för honungens antiinflammatoriska och sårläkande egenskaper (Stephens et al., 2010). Det bör noteras att inte alla Leptospermum-arter producerar aktiv honung, och även inom L. scoparium och L. polygalifolium-honung kan MGO-nivåerna variera från cirka 100 till över 1200 ppm (Windsor et al., 2012). En undersökning av australiensisk honungsaktivitet visade att honung från Leptospermum-växter som växer runt gränsen mellan New South Wales och Queensland var särskilt aktiv, men om detta beror på växt, jord, klimat eller andra faktorer är inte känt (Irish et al., 2011).
Hämning av patogener av honung
Honung har testats in vitro på en mängd olika patogener, särskilt de som kan kolonisera hud, sår och slemhinnor, där lokal behandling med honung är möjlig. Hittills har in vitro-tester visat att Manuka-honung effektivt kan hämma alla problematiska bakteriella patogener som testats (sammanfattat i Tabell 1). Särskilt intressant är att kliniska isolat med multiresistenta (MDR) fenotyper inte visar någon minskning i känslighet för honung, vilket tyder på en bred verkningsspektrum som skiljer sig från alla kända antimikrobiella medel (Willix et al., 1992; Blair och Carter, 2005; George och Cutting, 2007; Tan et al., 2009). Dessutom har försök att framställa honungsresistenta stammar i laboratoriet inte varit framgångsrika och det finns inga rapporter om kliniska isolat med förvärvad resistens mot honung (Blair et al., 2009; Cooper et al., 2010).
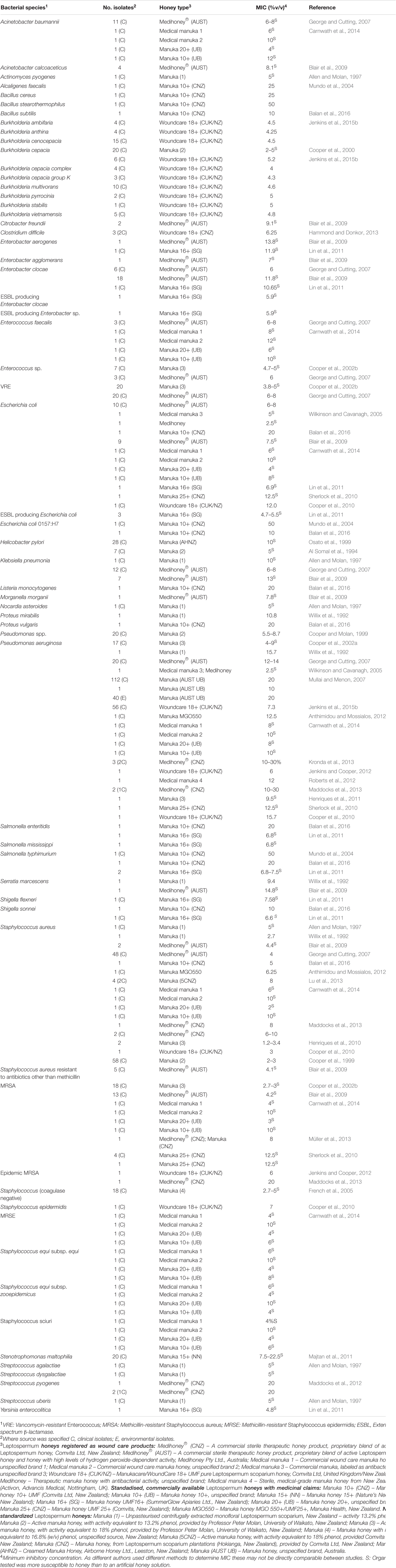
TABELL 1. Bakteriearter som visat sig vara känsliga för terapeutisk Manuka-honung.
Förutom att hämma planktoniska celler kan honung sprida och döda bakterier som lever i biofilmer. Biofilmer är cellgemenskaper som vanligtvis är inneslutna i en självproducerad extracellulär matris och finns fästa vid ytor, inklusive sår, tänder, slemhinnor och implanterade enheter. Mikrober som finns i biofilmer är skyddade från antimikrobiella medel och kan orsaka långvariga, icke-läkande infektioner. Manuka honung stör cellulära aggregat (Maddocks et al., 2012; Roberts et al., 2012) och förhindrar bildandet av biofilmer av ett brett spektrum av problematiska patogener, inklusive Streptococcus och Staphylococcus-arter, Pseudomonas aeruginosa, Escherichia coli, Proteus mirabilis, Enterobacter cloacae, Acinetobacter baumannii och Klebsiella pneumonia (Maddocks et al., 2012, 2013; Lu et al., 2014; Majtan et al., 2014a; Halstead et al., 2016). Viktigt är att honung också kan bryta ner etablerade biofilmer och döda de boende cellerna, även om en högre koncentration krävs än för planktoniska celler (Okhiria et al., 2009; Maddocks et al., 2013; Lu et al., 2014; Majtan et al., 2014a). Mycket nyligen testades manuka honung på en multispecies biofilm innehållande Staphylococcus aureus, Streptococcus agalactiae, Pseudomonas aeruginosa och Enterococcus faecalis och visade sig minska livskraften hos alla arter utom E. faecalis, som inte kunde utrotas (Sojka et al., 2016). Detta har tydliga kliniska implikationer för användning av honung på sår som innehåller biofilmer, och att förstå hur biofilmen möjliggör för E. faecalis att överleva när den normalt dödas av honung är ett viktigt och intressant område för framtida studier. MGO verkar vara mestadels men inte helt ansvarigt för hämningen av biofilmer av manuka honung, vilket återigen understryker vikten av ytterligare komponenter som modulerar aktiviteten (Kilty et al., 2011; Lu et al., 2014).
Honungens verkningsspektrum mot icke-bakteriella patogener är ännu inte väl fastställt. Nya studier som undersökt den antivirala effekten av Manuka-honung har föreslagit att den har potential för behandling av varicella-zosterviruset (orsaken till vattkoppor och bältros) (Shahzad och Cohrs, 2012) och influensa (Watanabe et al., 2014). Svampinfektioner i huden, inklusive Candida albicans och dermatofyter, är betydligt mindre känsliga än bakterier för Manuka-honung, men hämmas av honung med höga nivåer av väteperoxidproduktion (Brady et al., 1996; Irish et al., 2006). Manuka- och icke-Manuka-honung har visat sig minska livskraften hos sporer av mikrosporidianen Nosema apis, en viktig patogen hos bin, men honung kunde inte bota binens infektion när den väl påbörjats (Malone et al., 2001). Det har gjorts mycket få studier på användning av honung mot protozoer eller helminter, och dessa har inte använt honung med välkarakteriserad aktivitet, vilket gör det svårt att bedöma betydelsen av deras resultat (Bassam et al., 1997; Nilforoushzadeh et al., 2007; Sajid och Azim, 2012).
Att föra honung in i mainstreammedicin: Nya experimentella och mekanistiska studier belyser hur honung fungerar
Aktiv manukahonung finns allmänt tillgänglig som ett terapeutiskt medel och funktionellt livsmedel, och de flesta konsumenter accepterar det som en holistisk, något mystisk produkt. Brist på förståelse för hur honung dödar bakterier och främjar läkning begränsar dock dess acceptans inom konventionell medicin där det fortfarande betraktas som ”alternativt” eller ”komplementärt”. Majoriteten av forskningsstudier om honung hittills har varit beskrivande, men nyare studier försöker nu reda ut hur honung fungerar och använder mekanistiska metoder för att fastställa hur den verkar på cellulär och molekylär nivå.
Ultrastrukturella studier av bakterieceller och samhällen behandlade med honung
Honung kan kraftigt förändra storleken och formen på bakterieceller, även om omfattningen varierar mellan olika bakteriearter. Med hjälp av transmissions-elektronmikroskopi (TEM) hade S. aureus-kulturer behandlade med manukahonung fler celler med fullbordade septa jämfört med de som behandlades med artificiell honung, vilket tyder på att cellerna gick in i men misslyckades med att slutföra delningsstadiet i cellcykeln, även om dessa celler utåt sett såg normala ut vid svepelektronmikroskopi (SEM) (Henriques et al., 2010). Nyligen visade fas-kontrastavbildning efter behandling med en subletal dos manukahonung att celler av S. aureus och Bacillus subtilis var betydligt mindre och hade större sannolikhet att ha kondenserat DNA än de som växte utan honung (Lu et al., 2013). Det är svårt att direkt jämföra dessa studier eftersom de använde olika mängder honung och behandlingstider, men överlag tyder resultaten på en frånkoppling av tillväxt och celldelning, vilket ofta ses som svar på närings- och miljöstress (Silva-Rocha and de Lorenzo, 2010).
Honungsbehandling har rapporterats orsaka att kulturer av de gramnegativa arterna E. coli och P. aeruginosa har både onormalt korta och långa celler (Lu et al., 2013). Intressant nog verkar P. aeruginosa vara mindre mottaglig för hämning av honung än andra arter, men djupgående cellulära förändringar sågs med TEM och SEM, inklusive fåror och blåsor (utbuktningar av cellens plasmamembran) på cellens yta samt en betydande mängd extracellulärt skräp som tyder på celllys (Henriques et al., 2011). Detta bekräftades i en efterföljande studie med BacLight live-dead fluorescensfärgning och konfokalmikroskopi, även om denna också visade att ett relativt stort antal levande celler kvarstod. Dessa studier använde 20 % (v/v) honung, vilket var högre än MBC för deras stam av P. aeruginosa och betydande hämning och död förväntades. Dock fann atomkraftsmikroskopi (AFM) med sub-baktericida nivåer fortfarande betydande celldistorsion och blåsbildning i celler behandlade med MIC (12 %) och halva MIC (6 %) koncentrationer, tillsammans med omfattande celllys (Roberts et al., 2012). Denna uppenbara degeneration av P. aeruginosa-cellen stöddes av kvantitativ PCR-analys som visade en tiofaldig nedreglering i honungsbehandlade celler av oprF, som kodar för en yttre membranporin som är viktig för strukturell stabilitet (Jenkins et al., 2015a).
‘Omics-analyser bedömer hela cellens respons på hämning av honung
Möjligheten att bedöma hela cellers utsläpp har revolutionerat studiet av läkemedels-patogeninteraktioner och har särskilt värde för komplexa naturprodukter som honung där effekter på flera processer är sannolika. Microarray- och proteomstudier av bakterier exponerade för honung antydde en induktion av stressrelaterade processer och undertryckande av proteinsyntes (Blair et al., 2009; Jenkins et al., 2011; Packer et al., 2012). Även om detta övergripande är ganska typiskt för ett svar på hämmande medel, producerade honung ett unikt ”signatur” av differentierad uttryck som inkluderade många proteiner med hypotetiska eller okända funktioner, vilket tyder på en ny verkningsmekanism. Specifika gener eller proteiner som visade sig vara nedreglerade i ’omiks’-analyser av S. aureus och E. coli O157/H7 har funktioner relaterade till virulens, quorum sensing och biofilmsbildning (Lee et al., 2011; Jenkins et al., 2013), och i P. aeruginosa fanns en nedreglering av proteiner involverade i flagellering (Roberts et al., 2015). Dessa fenotyper är avgörande för att patogener ska kunna etablera sig och producera invasiv infektion och indikerar att honung, förutom att hämma tillväxt, kan minska den patogena potentialen hos infekterande bakterier.
Även om de fortfarande är relativt få och begränsade i omfattning, tyder de ’omiks’-analyser som genomförts hittills på ett komplext cellulärt svar på honung med betydande variation mellan olika bakteriearter. Avancerade systembiologiska metoder som möjliggör kontextualisering av data, samt valideringsstudier med kvantitativ PCR och genborttagningsstammar, krävs nu för att reda ut denna komplexitet, och dessa kan avslöja nya tillvägagångssätt för läkemedelsbehandlingar som syftar till att hämma bakterietillväxt (Hudson et al., 2012).
Interaktioner mellan honung och konventionella antibiotika
Förutom att användas som en ensam behandling finns det möjligheter att använda honung för att komplettera behandling med konventionella antibiotika. Detta kan vara särskilt värdefullt när det kombineras med systemiska medel som kan levereras till ett sår via blodcirkulationen medan honung appliceras topiskt. Kombinerade behandlingar kan också sänka den terapeutiska dosen av antimikrobiella medel och förhindra utveckling av resistens, och i vissa fall kan det leda till läkemedelssynergi, där den kombinerade aktiviteten är större än summan av de individuella aktiviteterna hos varje läkemedelspartner.
In vitro-studier som kombinerar terapeutiskt godkänd manuka honung med antibiotika har funnit en synergistisk effekt med oxacillin, tetracyklin, imipenem och mupirocin mot tillväxten av en MRSA-stam (Jenkins och Cooper, 2012). Dessutom återställde närvaron av en subinhibitorisk koncentration av honung i kombination med oxacillin MRSA-stammen till oxacillin-känslighet. Författarna fann nedreglering av mecR1, som kodar för ett MRSA-specifikt penicillinbindande protein (PBP2A) och föreslog detta som en mekanism för honungssynergi. Stark synergistisk aktivitet mellan manuka honung och rifampicin mot flera S. aureus-stammar, inklusive kliniska isolat och MRSA-stammar, har också påvisats, och närvaron av honung förhindrade uppkomsten av rifampicinresistens in vitro (Müller et al., 2013). Detta är kliniskt betydelsefullt eftersom rifampicin penetrerar väl in i vävnader och abscesser och ofta används för att behandla ytliga stafylokockinfektioner, men inducerar snabbt resistens och måste därför användas i kombination med ett annat medel. En ytterligare upptäckt från denna studie var att synergier inte berodde på MGO, eftersom en syntetisk honung tillsatt med MGO inte var synergistisk med rifampicin.
Att förstå hur honung påverkar verkan av antimikrobiella med välkarakteriserade verkningsmekanismer kan också fördjupa vår förståelse av hur honung påverkar bakteriella patogener. Liu et al. (2014) utvidgade analysen av synergier för att inkludera ytterligare antibiotika och olika S. aureus- och MRSA-stammar. De föreslog att en ökad känslighet för klindamycin och gentamicin kan bero på den kombinerade effekten av nedreglerad proteinsyntes av honung tillsammans med hämning av ribosomer av antibiotika, medan synergier med β-laktamantibiotika kan bero på ökad oxidativ stress orsakad av båda parterna. Eftersom S. aureus- och MRSA-stammar var lika känsliga för oxacillin-honungskombinationen verkade det osannolikt att synergier berodde på nedreglering av PBP2A. I en klinisk MRSA-isolat fanns dock ingen ökad känslighet för klindamycin eller gentamicin när honung var närvarande, vilket är anmärkningsvärt eftersom det är det första rapporterade fallet av en skillnad i respons på honung mellan MRSA och S. aureus. Att undersöka denna stam-specifika skillnad med hjälp av transkriptom- eller proteomanalyser skulle vara en intressant väg för framtida forskning (Liu et al., 2014).
Bevis för effektivitet från djurstudier, fallrapporter och kliniska prövningar
Företag som producerar och marknadsför manuka honung främjar höga etiska standarder och avråder från användning av djurmodeller för att studera infektioner och sårläkning. Manuka honung har dock använts för att behandla djur med kirurgiska eller oavsiktliga sår, särskilt hästar, med positiva resultat (Dart et al., 2015; Bischofberger et al., 2016). Fallrapporter som använder honung för icke-läkande sår och sårbildningar har noterat betydande förbättring med upplösning av infektion där konventionella antibiotika hade misslyckats (Regulski, 2008; Smith et al., 2009). Trots detta och bevis från många in vitro- och in vivo-modeller som visar att honung dödar problematiska sårpatogener, finns det en brist på robusta kliniska data för manuka honung. Det finns olika orsaker till detta, inklusive tekniska svårigheter att genomföra en dubbelblind placebokontrollerad studie på ett distinkt ämne som honung, etiska överväganden, bristande intresse från kliniska praktiker och kostnad kontra nytta för honungsföretag, vars fokus är på naturliga produkter och försäljning över disk där manuka honung och tillhörande förband redan har ett premiumpris. Dessa kan förändras i takt med att antibiotikaresistens urholkar nuvarande behandlingsalternativ och pågående forskning som lyfter fram honungens potential uppmärksammar medicinska praktiker.
Luckor och framväxande möjligheter i studiet av honung
Stora framsteg har nyligen gjorts i vår förståelse av terapeutisk honung, men dess användning inom klinisk medicin är fortfarande begränsad, även när konventionella antibiotika börjar misslyckas. Komplexiteten i honung, som kan anses vara dess största styrka i att döda olika patogener och förhindra resistens, försvårar dess studie eftersom många faktorer som verkar tillsammans sannolikt påverkar aktiviteten. Vi förespråkar ytterligare mekanistiska studier med lämpligt registrerad terapeutisk manuka honung, särskilt studier som använder icke-reduktionistiska systembiologiska metoder, tillsammans med detaljerade kemiska och mikrobiologiska analyser för att klargöra hur honung verkar på molekylär, cellulär och populationsnivå, hur detta kan skilja sig mellan olika stammar och arter av mikrobiella patogener, och hur värdcellen svarar (Tabell 2). Information som erhållits från dessa studier kan sedan informera terapin och producera de kliniska data som krävs för att ta honung in i mainstream-medicinen; inte längre en alternativ behandling som endast används när allt annat har misslyckats.
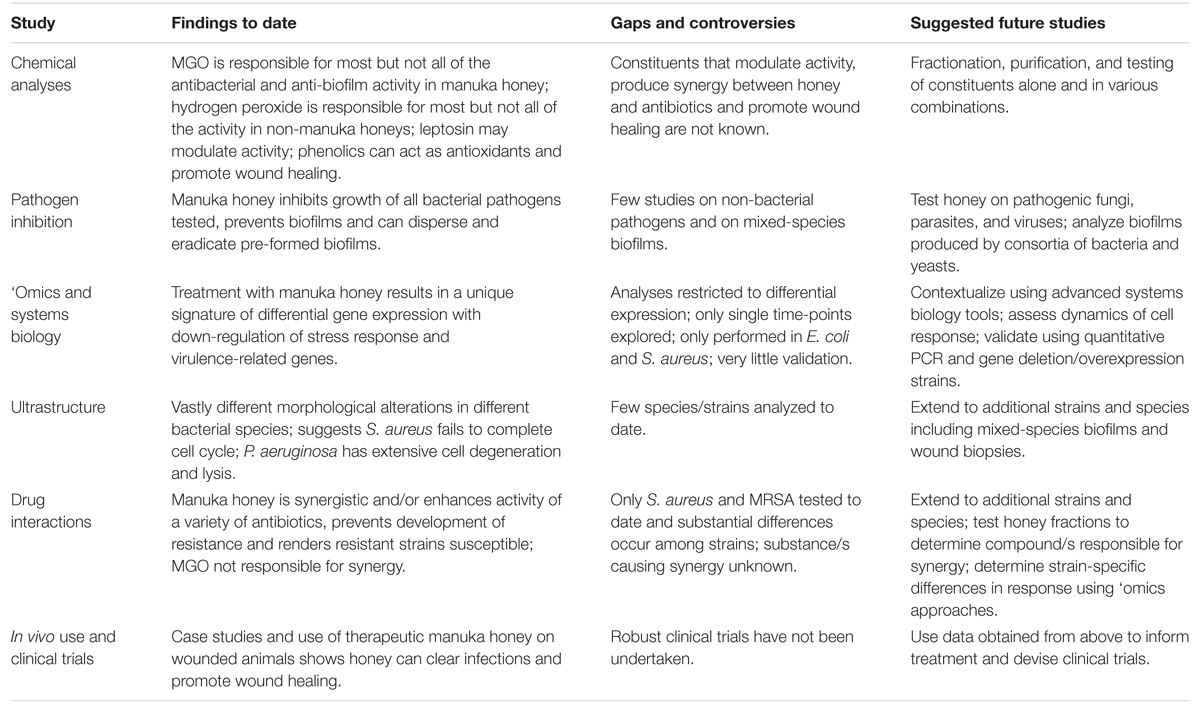
TABELL 2. Studier av manuka honung: fynd, luckor och framtida studier.
Författarbidrag
Denna översikt skrevs av DC, SB, NNC, DB och PB och granskades kritiskt av RS och EH.
Finansiering
NNC får lönebidrag från Rural Industries Research and Development Corporation – Honey Bee Program (Bidrag PRJ-009186).
Intressekonfliktdeklaration
DC, PB och EH rapporterar bidrag och icke-finansiellt stöd i form av manuka-honung från Comvita NZ Limited och Capilano Honey Limited; RS är anställd av Comvita NZ Limited, som handlar med medicinsk manuka-honung (Medihoney).
Övriga författare intygar att forskningen genomfördes utan någon kommersiell eller ekonomisk relation som kan tolkas som en potentiell intressekonflikt.
Förkortning
ESBL, utvidgat spektrum β-laktamas; MBC, minsta bakteriedödande koncentration; MGO, metylglyoxal; MIC, minsta hämmande koncentration; MRSA, meticillinresistent Staphylococcus aureus; MRSE, meticillinresistent Staphylococcus epidermis; NPA, icke-peroxidaktivitet; VRE, vankomycinresistent Enterococcus.
Referenser
Adams, C. J., Boult, C. H., Deadman, B. J., Farr, J. M., Grainger, M. N. C., Manley-Harris, M., et al. (2008). Isolering med HPLC och karaktärisering av den bioaktiva fraktionen av nyzeeländsk manuka (Leptospermum scoparium)-honung. Carbohydr. Res. 343, 651–659. doi: 10.1016/j.carres.2007.12.011
Adams, C. J., Manley-Harris, M., och Molan, P. C. (2009). Ursprung för metylglyoxal i nyzeeländsk manuka (Leptospermum scoparium)-honung. Carbohydr. Res. 344, 1050–1053. doi: 10.1016/j.carres.2009.03.020
Al Somal, N., Coley, K. E., Molan, P. C., och Hancock, B. M. (1994). Känslighet hos Helicobacter pylori för den antibakteriella aktiviteten hos manuka-honung. J. R. Soc. Med. 87, 9–12.
Allen, K., Molan, P., och Reid, G. (1991). En undersökning av den antibakteriella aktiviteten hos vissa honungssorter från Nya Zeeland. J. Pharm. Pharmacol.43, 817–822. doi: 10.1111/j.2042-7158.1991.tb03186.x
Allen, K. L., och Molan, P. C. (1997). Känsligheten hos mastitframkallande bakterier för honungens antibakteriella aktivitet. N. Z. J. Agric. Res. 40, 537–540. doi: 10.1080/00288233.1997.9513276
Anthimidou, E., och Mossialos, D. (2012). Antibakteriell aktivitet hos grekisk och cypriotisk honung mot Staphylococcus aureus och Pseudomonas aeruginosa jämfört med Manuka honung. J. Med. Food 16, 42–47. doi: 10.1089/jmf.2012.0042
Balan, P., Mal, G., Das, S., och Singh, H. (2016). Synergistiska och additiva antimikrobiella aktiviteter av curcumin, Manuka honung och vassleproteiner. J. Food Biochem. doi: 10.1111/jfbc.12249
Bassam, Z., Zohra, B. I., och Saada, A.-A. (1997). Effekterna av honung på Leishmania-parasiter: en in vitro-studie. Trop. Doctor27, 36–38.
Biglari, B., Moghaddam, A., Santos, K., Blaser, G., Büchler, A., Jansen, G., et al. (2013). Multicenter prospektiv observationsstudie om professionell sårvård med honung (Medihoney). Int. Wound J. 10, 252–259. doi: 10.1111/j.1742-481X.2012.00970.x
Bischofberger, A., Dart, C., Horadagoda, N., Perkins, N., Jeffcott, L., Little, C., et al. (2016). Effekt av Manuka honungsgel på koncentrationerna av transforming growth factor β1 och β3, bakterieantal och histomorfologi hos kontaminerade fulltjocklekssår i hästars distala ben. Aust. Vet. J. 94, 27–34. doi: 10.1111/avj.12405
Blair, S., Cokcetin, N., Harry, E., och Carter, D. (2009). Den ovanliga antibakteriella aktiviteten hos medicinsk Leptospermum-honung: antibakteriellt spektrum, resistens och transkriptomanalys. Eur. J. Clin. Microbiol. Infect. Dis. 28, 1199–1208. doi: 10.1007/s10096-009-0763-z
Blair, S. E., och Carter, D. A. (2005). Potentialen för honung vid hantering av sår och infektioner. J. Austral. Infect. Control 10, 24–31.
Brady, N., Molan, P., och Harfoot, C. (1996). Dermatofyters känslighet för den antimikrobiella aktiviteten hos manuka-honung och annan honung. Pharm. Pharmacol. Commun. 2, 471–473.
Carnwath, R., Graham, E. M., Reynolds, K., och Pollock, P. J. (2014). Honungens antimikrobiella aktivitet mot vanliga bakteriella isolat från häst sår. Vet. J. 199, 110–114. doi: 10.1016/j.tvjl.2013.07.003
Chen, C., Campbell, L., Blair, S. E., och Carter, D. A. (2012). Effekten av värmebehandling på honungens antimikrobiella egenskaper. Front. Microbiol. 3:265. doi: 10.3389/fmicb.2012.00265
Cooper, R., Jenkins, L., Henriques, A., Duggan, R., och Burton, N. (2010). Avsaknad av bakteriell resistens mot medicinsk manuka-honung. Eur. J. Clin. Microbiol. Infect. Dis. 29, 1237–1241. doi: 10.1007/s10096-010-0992-1
Cooper, R. A., Halas, E., och Molan, P. C. (2002a). Effektiviteten hos honung att hämma stammar av Pseudomonas aeruginosa från infekterade brännskador. J. Burn Care Rehabil. 23, 366–370. doi: 10.1097/00004630-200211000-00002
Cooper, R. A., och Molan, P. C. (1999). Användning av honung som antiseptikum vid behandling av Pseudomonas-infektion. J. Wound Care 8, 161–164. doi: 10.12968/jowc.1999.8.4.25867
Cooper, R. A., Molan, P. C., och Harding, K. G. (1999). Antibakteriell aktivitet hos honung mot stammar av Staphylococcus aureus från infekterade sår. J. R. Soc. Med. 92, 283–285.
Cooper, R. A., Molan, P. C., och Harding, K. G. (2002b). Känslighet för honung hos Gram-positiva kocker av klinisk betydelse isolerade från sår. J. Appl. Microbiol. 93, 857–863. doi: 10.1046/j.1365-2672.2002.01761.x
Cooper, R. A., Wigley, P., och Burton, N. F. (2000). Känslighet hos multiresistenta stammar av Burkholderia cepacia mot honung. Lett. Appl. Microbiol. 31, 20–24. doi: 10.1046/j.1472-765x.2000.00756.x
Dart, A., Bischofberger, A., Dart, C., och Jeffcott, L. (2015). En översikt av forskning om sekundärsårsläkning hos häst med manuka honung: nuvarande rekommendationer och framtida tillämpningar. Equine Vet. Educ. 27, 658–664. doi: 10.1111/eve.12379
Estevinho, L., Pereira, A. P., Moreira, L., Dias, L. G., och Pereira, E. (2008). Antioxidativa och antimikrobiella effekter av fenoliska föreningars extrakt från honung från nordöstra Portugal. Food Chem. Toxicol. 46, 3774–3779. doi: 10.1016/j.fct.2008.09.062
French, V. M., Cooper, R. A., och Molan, P. C. (2005). Den antibakteriella aktiviteten hos honung mot koagulasnegativa Staphylokocker. J. Antimicrobial Chemother. 56, 228–231. doi: 10.1093/jac/dki193
George, N. M., och Cutting, K. F. (2007). Antibakteriell honung (Medihoney): in-vitro-aktivitet mot kliniska isolat av MRSA, VRE och andra multiresistenta gramnegativa organismer inklusive Pseudomonas aeruginosa. Wounds 19:231.
Halstead, F. D., Webber, M. A., Rauf, M., Burt, R., Dryden, M., och Oppenheim, B. A. (2016). In vitro-aktivitet hos en konstruerad honung, medicinska honungssorter och antimikrobiella sårförband mot biofilmsproducerande kliniska bakterieisolat. J. Wound Care 25, 93–102. doi: 10.12968/jowc.2016.25.2.93
Hammond, E. N., och Donkor, E. S. (2013). Antibakteriell effekt av Manuka honung på Clostridium difficile. BMC Res. 6:188. doi: 10.1186/1756-0500-6-188
Henriques, A. F., Jenkins, R. E., Burton, N. F., och Cooper, R. A. (2010). De intracellulära effekterna av Manuka honung på Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 29, 45–50. doi: 10.1007/s10096-009-0817-2
Henriques, A. F., Jenkins, R. E., Burton, N. F., och Cooper, R. A. (2011). Effekten av Manuka honung på strukturen hos Pseudomonas aeruginosa. Eur. J. Clin. Microbiol. Infect. Dis. 30, 167–171. doi: 10.1007/s10096-010-1065-1
Hudson, N. J., Dalrymple, B. P., och Reverter, A. (2012). Bortom differentialuttryck: jakten på kausala mutationer och effektormolekyler. BMC Genomics 13:356. doi: 10.1186/1471-2164-13-356
Irish, J., Blair, S., och Carter, D. (2011). Den antibakteriella aktiviteten hos honung från australiensisk flora. PLoS ONE 6:e18229. doi: 10.1371/journal.pone.0018229
Irish, J., Carter, D. A., Shokohi, T., och Blair, S. E. (2006). Honung har en antifungal effekt mot Candida-arter. Med. Mycol. 44, 289–291. doi: 10.1080/13693780600931986
Jenkins, R., Burton, N., och Cooper, R. (2011). Effekten av manuka honung på uttrycket av universal stress protein A i meticillinresistent Staphylococcus aureus. Int. J. Antimicrob. Agents 37, 373–376. doi: 10.1016/j.ijantimicag.2010.11.036
Jenkins, R., Burton, N., och Cooper, R. (2013). Proteomisk och genomisk analys av meticillinresistent Staphylococcus aureus (MRSA) exponerad för manuka honung in vitro visade nedreglering av virulensmarkörer. J. Antimicrobial Chemother. 69, 603–615. doi: 10.1093/jac/dkt430
Jenkins, R., och Cooper, R. (2012). Förbättring av antibiotikans aktivitet mot sårpatogener med Manuka-honung in vitro. PLoS ONE 7:e45600. doi: 10.1371/journal.pone.0045600
Jenkins, R., Roberts, A., och Brown, H. L. (2015a). Om de antibakteriella effekterna av Manuka-honung: mekanistiska insikter. Res. Rep. Biol. 6, 215–224. doi: 10.2147/RRB.S75754
Jenkins, R., Wootton, M., Howe, R., och Cooper, R. (2015b). En demonstration av känsligheten hos kliniska isolat från cystisk fibros-patienter för Manuka-honung. Arch. Microbiol. 197, 597–601. doi: 10.1007/s00203-015-1091-6
Kalapos, M. P. (2008). Tandemreaktionen mellan fria radikaler och methylglyoxal. Chem. Biol. Interact. 171, 251–271. doi: 10.1016/j.cbi.2007.11.009
Kato, Y., Umeda, N., Maeda, A., Matsumoto, D., Kitamoto, N., och Kikuzaki, H. (2012). Identifiering av en ny glycosid, leptosin, som en kemisk markör för Manuka-honung. J. Agric. Food Chem. 60, 3418–3423. doi: 10.1021/jf300068w
Kilty, S. J., Duval, M., Chan, F. T., Ferris, W., och Slinger, R. (2011). Methylglyoxal: (aktivt ämne i Manuka-honung) in vitro-aktivitet mot bakteriella biofilmer. Int. Forum Allergy Rhinol. 1, 348–350. doi: 10.1002/alr.20073
Kronda, J. M., Cooper, R. A., och Maddocks, S. E. (2013). Manuka-honung hämmar produktionen av sideroforer i Pseudomonas aeruginosa. J. Appl. Microbiol. 115, 86–90. doi: 10.1111/jam.12222
Kwakman, P. H., te Velde, A. A., de Boer, L., Vandenbroucke-Grauls, C. M., och Zaat, S. A. (2011). Två stora medicinska honungssorter har olika mekanismer för bakteriedödande aktivitet. PLoS ONE 6:e17709. doi: 10.1371/journal.pone.0017709
Lee, J.-H., Park, J.-H., Kim, J.-A., Neupane, G. P., Cho, M. H., Lee, C.-S., et al. (2011). Låga koncentrationer av honung minskar biofilmsbildning, quorum sensing och virulens hos Escherichia coli O157: H7. Biofouling 27, 1095–1104. doi: 10.1080/08927014.2011.633704
Lin, S. M., Molan, P. C., och Cursons, R. T. (2011). Den kontrollerade in vitro känsligheten hos gastrointestinala patogener för den antibakteriella effekten av Manuka-honung. Eur. J. Clin. Microbiol. Infect. Dis. 30, 569–574. doi: 10.1007/s10096-010-1121-x
Liu, M., Lu, J., Müller, P., Turnbull, L., Burke, C. M., Schlothauer, R. C., et al. (2014). Antibiotikaspecifika skillnader i svaret från Staphylococcus aureus vid behandling med antimikrobiella medel kombinerade med Manuka-honung. Front. Microbiol. 5:779. doi: 10.3389/fmicb.2014.00779
Lu, J., Carter, D. A., Turnbull, L., Rosendale, D., Hedderley, D., Stephens, J., et al. (2013). Effekten av nyzeeländsk kanuka, manuka och klöverhonung på bakteriell tillväxtdynamik och cellmorfologi varierar beroende på art. PLoS ONE8:e55898. doi: 10.1371/journal.pone.0055898
Lu, J., Turnbull, L., Burke, C. M., Liu, M., Carter, D. A., Schlothauer, R. C., et al. (2014). Manuka-typ honung kan utrota biofilmer producerade av Staphylococcus aureus stammar med olika förmåga att bilda biofilm. PeerJ 2:e326. doi: 10.7717/peerj.326
Maddocks, S. E., Jenkins, R. E., Rowlands, R. S., Purdy, K. J., och Cooper, R. A. (2013). Manuka honung hämmar adhesion och invasion av medicinskt viktiga sårbakterier in vitro. Fut. Microbiol. 8, 1523–1536. doi: 10.2217/fmb.13.126
Maddocks, S. E., Lopez, M. S., Rowlands, R. S., och Cooper, R. A. (2012). Manuka honung hämmar utvecklingen av Streptococcus pyogenes biofilmer och orsakar minskad uttryck av två fibronektinbindande proteiner. Microbiology 158, 781–790. doi: 10.1099/mic.0.053959-0
Majtan, J. (2014). Honung: en immunmodulator vid sårläkning. Wound Repair Regenerat. 22, 187–192. doi: 10.1111/wrr.12117
Majtan, J., Bohova, J., Horniackova, M., Klaudiny, J., och Majtan, V. (2014a). Anti-biofilm-effekter av honung mot sårpatogenerna Proteus mirabilis och Enterobacter cloacae. Phytother. Res. 28, 69–75. doi: 10.1002/ptr.4957
Majtan, J., Bohova, J., Prochazka, E., och Klaudiny, J. (2014b). Methylglyoxal kan påverka ackumuleringen av väteperoxid i manuka-honung genom hämning av glukosoxidas. J. Med. Food 17, 290–293. doi: 10.1089/jmf.2012.0201
Majtan, J., Klaudiny, J., Bohova, J., Kohutova, L., Dzurova, M., Sediva, M., et al. (2012). Methylglyoxal-inducerade modifieringar av viktiga proteinkomponenter i honungsbin i manuka-honung: möjliga terapeutiska implikationer. Fitoterapia 83, 671–677. doi: 10.1016/j.fitote.2012.02.002
Majtan, J., Majtanova, L., Bohova, J., och Majtan, V. (2011). Honungsdaggshonung som en potent antibakteriell agent vid utrotning av multiresistenta Stenotrophomonas maltophilia-isolat från cancerpatienter. Phytother. Res. 25, 584–587. doi: 10.1002/ptr.3304
Malone, L. A., Gatehouse, H. S., och Tregidga, E. L. (2001). Effekter av tid, temperatur och honung på Nosema apis (Microsporidia: Nosematidae), en parasit hos honungsbiet, Apis mellifera (Hymenoptera: Apidae). J. Invertebrate Pathol. 77, 258–268. doi: 10.1006/jipa.2001.5028
Mavric, E., Wittmann, S., Barth, G., och Henle, T. (2008). Identifiering och kvantifiering av methylglyoxal som den dominerande antibakteriella beståndsdelen i Manuka (Leptospermum scoparium)-honung från Nya Zeeland. Mol. Nutrit. Food Res. 52, 483–489. doi: 10.1002/mnfr.200700282
Molan, P. M. (2008). En förklaring till varför MGO-nivån i Manuka-honung inte visar den antibakteriella aktiviteten. New Zealand Beekeeper 16, 11–13.
Mullai, V., och Menon, T. (2007). Baktericid aktivitet hos olika typer av honung mot kliniska och miljömässiga isolat av Pseudomonas aeruginosa. J. Alternat. Complement. Med. 13, 439–442. doi: 10.1089/acm.2007.6366
Müller, P., Alber, D. G., Turnbull, L., Schlothauer, R. C., Carter, D. A., Whitchurch, C. B., et al. (2013). Synergism mellan Medihoney och rifampicin mot meticillinresistent Staphylococcus aureus (MRSA). PLoS ONE 8:e57679. doi: 10.1371/journal.pone.0057679
Mundo, M. A., Padilla-Zakour, O. I., och Worobo, R. W. (2004). Tillväxthämning av livsmedelsburna patogener och livsmedelsförstörande organismer av utvalda råa honungssorter. Int. J. Food Microbiol. 97, 1–8. doi: 10.1016/j.ijfoodmicro.2004.03.025
Nilforoushzadeh, M. A., Jaffary, F., Moradi, S., Derakhshan, R., och Haftbaradaran, E. (2007). Effekt av topikal honungsapplicering tillsammans med intralesionell injektion av glucantime vid behandling av kutan leishmaniasis. BMC Complement Altern. Med. 7:1. doi: 10.1186/1472-6882-7-1
Norton, A. M., McKenzie, L. N., Brooks, P. R., och Pappalardo, L. J. (2015). Kvantifiering av dihydroxyaceton i australiensisk Leptospermum-nektar med hjälp av högpresterande vätskekromatografi. J. Agric. Food Chem. 63, 6513–6517. doi: 10.1021/acs.jafc.5b01930
Okhiria, O., Henriques, A., Burton, N., Peters, A., och Cooper, R. (2009). Honung påverkar biofilmer av Pseudomonas aeruginosa på ett tids- och dosberoende sätt. J. ApiProduct. ApiMedical Sci. 1, 6–10. doi: 10.3896/IBRA.4.01.1.03
Osato, M. S., Reddy, S. G., och Graham, D. Y. (1999). Osmotisk effekt av honung på tillväxt och livskraft hos Helicobacter pylori. Dig. Dis. Sci. 44, 462–464. doi: 10.1023/A:1026676517213
Packer, J. M., Irish, J., Herbert, B. R., Hill, C., Padula, M., Blair, S. E., et al. (2012). Specifik icke-peroxid antibakteriell effekt av Manuka-honung på Staphylococcus aureus-proteomet. Int. J. Antimicrob. Agents 40, 43–50. doi: 10.1016/j.ijantimicag.2012.03.012
Regulski, M. (2008). En ny förbandstyp för behandling av kroniska bensår. Podiatry Manag. 27, 235–246.
Roberts, A. E., Maddocks, S. E., och Cooper, R. A. (2012). Manuka-honung är bakteriedödande mot Pseudomonas aeruginosa och leder till differentierad uttryck av oprF och algD. Microbiology 158, 3005–3013. doi: 10.1099/mic.0.062794-0
Roberts, A. E. L., Maddocks, S. E., och Cooper, R. A. (2015). Manuka-honung minskar rörligheten hos Pseudomonas aeruginosa genom att undertrycka flagellassocierade gener. J. Antimicrob. Chemother. 70, 716–725. doi: 10.1093/jac/dku448
Sajid, M., och Azim, M. K. (2012). Karakterisering av den nematicida aktiviteten hos naturlig honung. J. Agric. Food Chem. 60, 7428–7434. doi: 10.1021/jf301653n
Shahzad, A., och Cohrs, R. J. (2012). In vitro antiviral aktivitet hos honung mot varicella zoster-virus (VZV): en translational medicinstudie för potentiell behandling av bältros. Transl. Biomed. 3:2.
Sherlock, O., Dolan, A., Athman, R., Power, A., Gethin, G., Cowman, S., et al. (2010). Jämförelse av den antimikrobiella aktiviteten hos Ulmo-honung från Chile och Manuka-honung mot meticillinresistent Staphylococcus aureus, Escherichia coli och Pseudomonas aeruginosa. BMC Complement Altern. Med. 10:47. doi: 10.1186/1472-6882-10-47
Silva-Rocha, R., och de Lorenzo, V. (2010). Brus och robusthet i prokaryota regleringsnätverk. Annu. Rev. Microbiol. 64, 257–275. doi: 10.1146/annurev.micro.091208.073229
Smith, T., Hanft, J. R., och Legel, K. (2009). Topikal Leptospermum-honung vid svårbehandlade venösa bensår: en preliminär fallserie. Adv. Skin Wound Care 22, 68–71. doi: 10.1097/01.ASW.0000345283.05532.9a
Soffer, A. (1976). Chihuahuor och laetrile, kelaterapi och honung från Boulder, Colorado [editorial]. Arch. Intern. Med.136, 865–866. doi: 10.1001/archinte.136.8.865
Sojka, M., Valachova, I., Bucekova, M., och Majtan, J. (2016). Antibiofilm-effektivitet hos honung och bi-deriverad defensin-1 på mångartade sårbiofilmer. J. Med. Microbiol. doi: 10.1099/jmm.0.000227 [Epub ahead of print].
Stephens, J. M., Schlothauer, R. C., Morris, B. D., Yang, D., Fearnley, L., Greenwood, D. R., et al. (2010). Fenoliska föreningar och metylglyoxal i några nyzeeländska Manuka- och Kanuka-honungssorter. Food Chem. 120, 78–86. doi: 10.1016/j.foodchem.2009.09.074
Tan, H. T., Rahman, R. A., Gan, S. H., Halim, A. S., Hassan, S. A., Sulaiman, S. A., et al. (2009). De antibakteriella egenskaperna hos malaysisk tualanghonung mot sår- och enteriska mikroorganismer jämfört med Manuka-honung. BMC Complement Altern. Med. 9:34. doi: 10.1186/1472-6882-9-34
Watanabe, K., Rahmasari, R., Matsunaga, A., Haruyama, T., och Kobayashi, N. (2014). Antiinfluensavirus-effekter av honung in vitro: stark hög aktivitet hos manuka-honung. Arch. Med. Res. 45, 359–365. doi: 10.1016/j.arcmed.2014.05.006
Wilkinson, J. M., och Cavanagh, H. M. A. (2005). Antibakteriell aktivitet hos 13 honungssorter mot Escherichia coli och Pseudomonas aeruginosa. J. Med. Food 8, 100–103. doi: 10.1089/jmf.2005.8.100
Williams, S., King, J., Revell, M., Manley-Harris, M., Balks, M., Janusch, F., et al. (2014). Regionala, årliga och individuella variationer i dihydroxyacetoninnehållet i nektar från manuka (Leptospermum scoparium) i Nya Zeeland. J. Agric. Food Chem. 62, 10332–10340. doi: 10.1021/jf5045958
Willix, D. J., Molan, P. C., och Harfoot, C. G. (1992). En jämförelse av känsligheten hos bakterier som infekterar sår för den antibakteriella aktiviteten hos manuka-honung och annan honung. J. Appl. Bacteriol. 73, 388–394. doi: 10.1111/j.1365-2672.1992.tb04993.x
Windsor, S., Pappalardo, M., Brooks, P., Williams, S., och Manley-Harris, M. (2012). En praktisk ny analys av dihydroxyaceton och methylglyoxal tillämpad på australiensiska Leptospermum-honungar. J. Pharmacogn. Phytother. 4, 6–11.
Terapeutisk Manuka-honung: Inte längre så alternativ - https://www.frontiersin.org/articles/10.3389/fmicb.2016.00569/full
Leave a comment