治療用Manukaハニー:もはや代替ではない

医療用はちみつの研究は大きな再興期を迎えています。かつては主流医療から「代替療法」としてほとんど無視されていた民間療法の一つでしたが、現在では科学者、臨床医、一般の人々の間ではちみつの治療利用に対する関心が高まっています。この関心の背景にはいくつかの要因があります。第一に、多くの細菌病原体による抗生物質耐性の増加が新しい抗菌剤の開発と使用への関心を促していること。第二に、信頼できる研究や症例報告が増え、特定のはちみつが非常に効果的な創傷治療に役立つことが示されていること。第三に、治療用はちみつは高価格で取引されており、はちみつ業界がこれを活用するための研究を積極的に推進していること。そして最後に、はちみつの非常に複雑で予測困難な性質が研究者にとって魅力的な挑戦となっていることです。本稿では、マヌカはちみつの研究について、その抗菌効果に関する観察研究から、はちみつを主流医療に取り入れることを目指した現在の実験的・機構的研究までを概観します。はちみつの作用機序に関する現在の知識のギャップや未解決の論争点を整理し、はちみつがもはや「代替」ではない代替療法となるための新たな研究の方向性を提案します。
はじめに
ハチミツは人類の歴史を通じて薬として使用されてきました。ハチミツの最も一般的で持続的な治療用途の一つは創傷被覆材としてであり、これはほぼ間違いなくその抗菌特性によるものです。1960年代に高活性の抗生物質が登場すると、ハチミツは「無価値だが無害な物質」として軽視されました(Soffer, 1976)。しかし、現在進行中の抗生物質耐性の危機により、ハチミツはそれ自体が効果的な治療剤として、また新しい治療法を開発するための治療的手がかりとして再び注目されています。ハチミツは通常、花の蜜から採取され、主にヨーロッパミツバチApis melliferaによって生産される、糖類、アミノ酸、フェノール類、その他の物質の複雑な混合物です。異なる花から採取されたハチミツの種類は細菌を殺す能力に大きな差があり、これがハチミツに関する文献を複雑にし、異なる研究間で結果を再現することを時に困難にしています(Allen et al., 1991; Irish et al., 2011)。最近の研究の大部分は、ニュージーランドとオーストラリア原産の特定のLeptospermum種から生産され、適切な医療規制機関に創傷ケア製品として登録されている、よく特徴付けられ標準化された活性マヌカハニーの作用機序に焦点を当てています。したがって、特に指定がない限り、本レビューはマヌカハニーに焦点を当てます。
活性マヌカハニーの化学分析
ニュージーランドのワイカト大学のピーター・モラン教授は、1980年代半ばにマヌカハニーの特異な活性を初めて報告し、さまざまな細菌種に対するその作用を試験し始めました。しかし、マヌカハニーの低濃度でも細菌病原体を殺菌することは明らかでしたが、その特定の活性成分は長年にわたり不明のままでした。高い糖度と低いpHは微生物の成長を抑制しますが、これらが希釈されて無視できるレベルになっても活性は残ります。多くの異なる種類のはちみつも、ミツバチ由来のグルコースオキシダーゼがグルコースと水と反応して過酸化水素を生成します。しかし、マヌカハニーでは過酸化水素の生成は比較的低く、カタラーゼによって中和されることもありますが、それでも活性は残ります。この残る活性は「非過酸化水素活性」またはNPAと呼ばれ、2008年に2つの研究所が独立してマヌカハニー中のメチルグリオキサール(MGO)を特定したことでついに解明されました(Adams et al., 2008; Mavric et al., 2008)。MGOは、その前駆体であるジヒドロキシアセトン(DHA)が自然に脱水されて生成されるもので、これはニュージーランドとオーストラリア原産のLeptospermum scoparium、Leptospermum polygalifolium、および関連するLeptospermum種の花の蜜に含まれる天然の植物化学物質です(Adams et al., 2009; Williams et al., 2014; Norton et al., 2015)。MGOはDNA、RNA、タンパク質などの高分子と比較的非特異的に反応することができ(Adams et al., 2008; Mavric et al., 2008; Majtan et al., 2014b)、理論的には哺乳類細胞に対しても毒性を持つ可能性があります(Kalapos, 2008)。しかし、マヌカハニーを経口摂取したり創傷被覆材として使用した場合に宿主細胞への損傷の証拠はなく、むしろ創傷に塗布すると治癒を促進し瘢痕を減少させることが示されています(Biglari et al., 2013; Majtan, 2014; Dart et al., 2015)。このように細菌細胞に対して選択的な毒性を示すメカニズムはまだ解明されていません。
MGOや過酸化水素の高レベルは通常、最も活性の高い蜂蜜を生み出しますが、その相関は必ずしも完全ではなく、蜂蜜の他の成分が活性を調節している可能性があります(Molan, 2008; Kwakman et al., 2011; Chen et al., 2012; Lu et al., 2013)。抗菌性のあるミツバチ由来ペプチドであるBee defensin-1は、出所不明の活性蜂蜜であるRevamil蜂蜜の活性に関与していますが、これはManuka蜂蜜では構造的に変化し、活性を示さないようです(Kwakman et al., 2011; Majtan et al., 2012)。LeptosinというグリコシドはLeptospermum蜂蜜にのみ含まれ、そのレベルは効力と相関し、Manuka蜂蜜の抗菌活性を調節している可能性があります(Kato et al., 2012)。同様に、特に色の濃い蜂蜜には抗菌活性の可能性がある様々なフェノール化合物が含まれており、これらは単独では抑制的なレベルではないものの、互いにまたは蜂蜜の他の成分と相乗効果を発揮して活性を生み出したり変化させたりすることがあります(Estevinho et al., 2008; Stephens et al., 2010)。フェノール類は抗酸化作用も持ち、蜂蜜の抗炎症および創傷治癒特性の原因となっている可能性もあります(Stephens et al., 2010)。すべてのLeptospermum種が活性蜂蜜を生産するわけではなく、L. scopariumやL. polygalifoliumの蜂蜜でもMGOレベルは約100から1200 ppm以上まで幅があります(Windsor et al., 2012)。オーストラリアの蜂蜜の活性調査では、ニューサウスウェールズ州とクイーンズランド州の境界付近で育つLeptospermum植物由来の蜂蜜が特に活性が高いことが分かりましたが、これが植物、土壌、気候、その他の要因によるものかは不明です(Irish et al., 2011)。
ハニーによる病原体の抑制
ハニーは、特に皮膚、創傷、粘膜に定着する可能性のある多様な病原体に対してin vitroで試験されており、局所的なハニー治療が可能な場所で効果が検証されています。これまでのin vitro試験では、マヌカハニーが試験されたすべての問題のある細菌病原体を効果的に抑制できることが示されています(表1にまとめられています)。特に注目すべきは、多剤耐性(MDR)表現型を持つ臨床分離株でもハニーに対する感受性が低下しないことであり、これは既知のどの抗菌剤とも異なる広範囲の作用を示しています(Willix et al., 1992;Blair and Carter, 2005;George and Cutting, 2007;Tan et al., 2009)。さらに、実験室でハニー耐性株を作り出そうとする試みは成功しておらず、ハニーに対する獲得耐性を持つ臨床分離株の報告もありません(Blair et al., 2009;Cooper et al., 2010)。
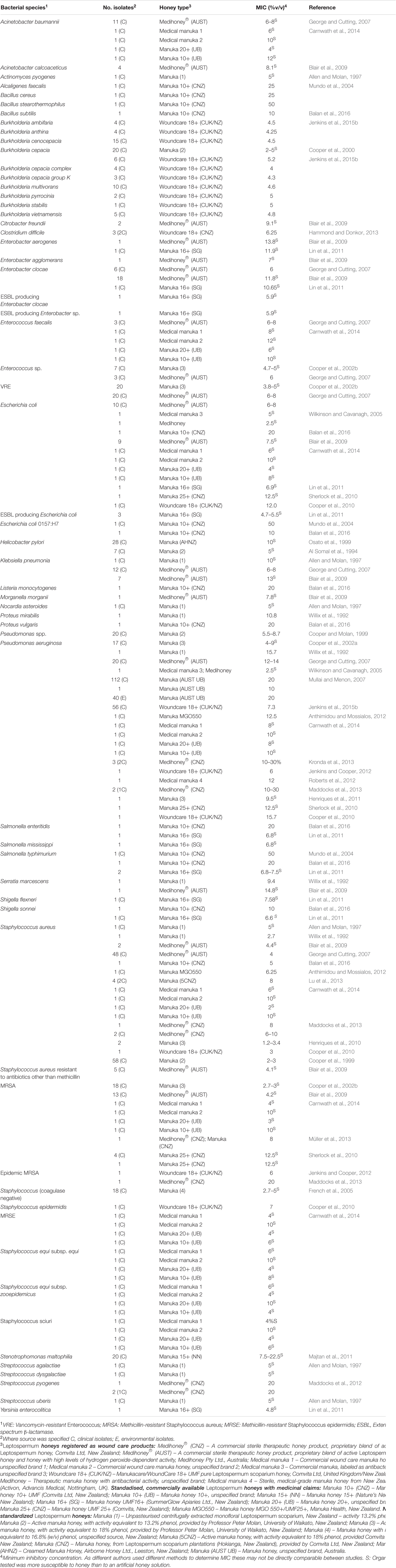
表1. 治療用マヌカハニーに感受性があると判明した細菌種。
プランクトン細胞の抑制に加え、はちみつはバイオフィルム内に生息する細菌を分散させ、殺菌することができます。バイオフィルムは、一般的に自己生成した細胞外マトリックスに包まれ、創傷、歯、粘膜表面、埋め込みデバイスなどの表面に付着している細胞の集合体です。バイオフィルム内に存在する微生物は抗菌剤から保護されており、持続的で解決しない感染症を引き起こすことがあります。Manukaは細胞の集合体を破壊し(Maddocks et al., 2012; Roberts et al., 2012)、StreptococcusやStaphylococcus種、Pseudomonas aeruginosa, Escherichia coli, Proteus mirabilis, Enterobacter cloacae, Acinetobacter baumannii、およびKlebsiella pneumoniaを含む幅広い問題のある病原体によるバイオフィルムの形成を防ぎます(Maddocks et al., 2012, 2013; Lu et al., 2014; Majtan et al., 2014a; Halstead et al., 2016)。重要なことに、はちみつは既に形成されたバイオフィルムを破壊し、そこに存在する細胞を殺すこともできますが、プランクトン細胞よりも高い濃度が必要です(Okhiria et al., 2009; Maddocks et al., 2013; Lu et al., 2014; Majtan et al., 2014a)。ごく最近、ManukaはStaphylococcus aureus, Streptococcus agalactiae, Pseudomonas aeruginosa、およびEnterococcus faecalisを含む多種バイオフィルムで試験され、すべての種の生存率を減少させましたが、通常ははちみつで殺されるE. faecalisは根絶できませんでした(Sojka et al., 2016)。これはバイオフィルムを含む創傷に対するはちみつの使用に明確な臨床的意義があり、バイオフィルムが通常はちみつで殺されるE. faecalisの生存を可能にする仕組みを理解することは、今後の重要かつ興味深い研究分野です。MGOはManukaによるバイオフィルム抑制の主な要因であるものの完全ではなく、活性を調節する追加成分の重要性を改めて示しています(Kilty et al., 2011; Lu et al., 2014)。
蜂蜜の非細菌性病原体に対する活性スペクトルはまだ十分に確立されていません。Manuka蜂蜜の抗ウイルス効果を調べた最近の研究では、水痘帯状疱疹ウイルス(水痘および帯状疱疹の原因)の治療に可能性が示唆されています(Shahzad and Cohrs, 2012)およびインフルエンザ(Watanabe et al., 2014)。皮膚の真菌病原体であるCandida albicansや皮膚糸状菌種は、Manuka蜂蜜に対して細菌よりもかなり感受性が低いですが、高レベルの過酸化水素生成を伴う蜂蜜によって抑制されます(Brady et al., 1996; Irish et al., 2006)。Manukaおよび非Manuka蜂蜜は、ミツバチの重要な病原体である微胞子虫Nosema apisの胞子の生存率を低下させることがわかっていますが、感染が進行した後のミツバチの感染を治癒することはできませんでした(Malone et al., 2001)。原虫や線虫寄生虫に対する蜂蜜の使用に関する研究は非常に少なく、これらの研究では活性が十分に特性評価された蜂蜜を使用していないため、その結果の重要性を評価することは困難です(Bassam et al., 1997; Nilforoushzadeh et al., 2007; Sajid and Azim, 2012)。
蜂蜜を主流医療に取り入れる:最近の実験的および機構的研究が蜂蜜の作用メカニズムを解明
活性マヌカハニーは治療剤や機能性食品として広く利用されており、多くの消費者はそれを全体的でやや神秘的な製品として受け入れています。しかし、蜂蜜がどのように細菌を殺し、治癒を促進するのかについての理解不足が、主流医療における受け入れを制限しており、依然として「代替」または「補完的」と見なされています。これまでの蜂蜜に関する研究の大部分は記述的でしたが、最近の研究では蜂蜜の作用機序を解明し、細胞および分子レベルでの作用を明らかにするための機構的アプローチが試みられています。
蜂蜜で処理された細菌細胞および集団の超微細構造研究
蜂蜜は細菌の大きさや形状に深い影響を与えることがありますが、その程度は細菌の種類によって異なります。透過型電子顕微鏡(TEM)を用いた研究では、マヌカハニーで処理されたS. aureus培養物は人工蜂蜜で処理されたものよりも完成した隔壁を持つ細胞が多く、細胞は細胞周期の分裂段階に入ったものの完了できなかったことを示唆していますが、走査型電子顕微鏡(SEM)ではこれらの細胞は外見上は正常に見えました(Henriques et al., 2010)。より最近の研究では、致死量未満のマヌカハニー処理後の位相差顕微鏡観察により、S. aureusおよびBacillus subtilisの細胞は有意に小さく、ハニーなしで成長した細胞よりもDNAが凝縮している可能性が高いことがわかりました(Lu et al., 2013)。これらの研究は使用したハニーの量や処理時間が異なるため直接比較は難しいですが、全体としては成長と細胞分裂の連結が外れた状態を示しており、これは栄養や環境ストレスに応答してよく見られる現象です(Silva-Rocha and de Lorenzo, 2010)。
ハチミツ処理は、グラム陰性菌のE. coliおよびP. aeruginosaの培養において、異常に短い細胞と長い細胞の両方を引き起こすことが報告されています(Lu et al., 2013)。興味深いことに、P. aeruginosaは他の種よりもハチミツによる阻害に対して感受性が低いように見えますが、TEMおよびSEMを用いた観察では、細胞表面に溝や膨らみ(細胞膜の突出)が見られ、細胞溶解を示す大量の細胞外破片が確認されました(Henriques et al., 2011)。これは、BacLight生死蛍光染色と共焦点顕微鏡を用いた後続の研究でも確認されましたが、比較的多くの生細胞が残存していることも示されました。これらの研究では20%(w/v)のハチミツを使用しており、これは彼らのP. aeruginosa株のMBCより高く、かなりの阻害と死滅が予想されます。しかし、亜殺菌レベルを用いた原子間力顕微鏡(AFM)では、MIC(12%)および半MIC(6%)濃度で処理された細胞においても、著しい細胞の変形と膨らみ、さらに細胞溶解が認められました(Roberts et al., 2012)。このP. aeruginosa細胞の明らかな変性は、構造的安定性に重要な外膜ポリンをコードするoprFのハチミツ処理細胞における10倍の発現低下を示した定量PCR解析によっても支持されました(Jenkins et al., 2015a)。
オミクス解析はハチミツによる阻害に対する全細胞応答を評価する
全細胞のアウトプットを評価する能力は、薬剤と病原体の相互作用の研究に革命をもたらし、蜂蜜のように複数のプロセスに影響を与える複雑な天然物に特に価値があります。蜂蜜に曝露された細菌のマイクロアレイおよびプロテオミクス研究は、ストレス関連プロセスの誘導とタンパク質合成の抑制を示唆しました(Blair et al., 2009; Jenkins et al., 2011; Packer et al., 2012)。全体としてこれは抑制剤に対する典型的な応答ですが、蜂蜜は多くの仮説的または未知の機能を持つタンパク質を含む独特の「シグネチャー」を生み出し、新しい作用機序を示唆しています。‘オミクス’解析で発現が抑制されたS. aureusおよびE. coli O157/H7の特定の遺伝子やタンパク質は、病原性、クオラムセンシング、バイオフィルム形成に関連する機能を持ち(Lee et al., 2011; Jenkins et al., 2013)、P. aeruginosaでは鞭毛形成に関与するタンパク質の発現抑制が見られました(Roberts et al., 2015)。これらの表現型は病原体が感染を確立し侵襲性の感染を引き起こすために重要であり、蜂蜜は増殖を抑制するだけでなく、感染細菌の病原性を低減できることを示しています。
これまでに行われた‘オミクス’解析はまだ数も範囲も限られていますが、蜂蜜に対する複雑な細胞応答が示唆されており、細菌種によって大きな違いがあります。データの文脈化を可能にする高度なシステム生物学的アプローチや、定量的PCRや遺伝子欠失株を用いた検証研究が今後必要であり、これにより細菌の増殖を抑制する薬物療法の新たな手法が明らかになる可能性があります(Hudson et al., 2012)。
蜂蜜と従来の抗生物質の相互作用
単独の治療剤としての使用に加え、蜂蜜は従来の抗生物質治療を補強するためにも利用可能です。これは、蜂蜜を局所的に適用しながら血液循環を通じて創傷床に届けられる全身性薬剤と組み合わせる場合に特に有用です。併用治療は抗菌剤の治療用量を減らし、耐性の発生を防ぐことができ、場合によっては薬剤の相乗効果をもたらし、各薬剤の個別の効果の合計を超える効果を生み出すこともあります。
in vitro研究では、治療的に承認されたManuka蜂蜜と抗生物質を組み合わせることで、MRSA株の増殖に対してオキサシリン、テトラサイクリン、イミペネム、ムピロシンとの相乗効果が見られました(Jenkins and Cooper, 2012)。さらに、蜂蜜のサブ阻害濃度がオキサシリンと組み合わさることで、MRSA株のオキサシリン感受性が回復しました。著者らは、MRSA特異的なペニシリン結合タンパク質(PBP2A)をコードするmecR1のダウンレギュレーションを発見し、これを蜂蜜の相乗効果のメカニズムとして示唆しました。Manuka蜂蜜とリファンピシンの間に、臨床分離株やMRSA株を含む複数のS. aureus株に対して強い相乗活性が認められ、蜂蜜の存在によりin vitroでリファンピシン耐性の出現が防がれました(Müllerら、2013年)。これは臨床的に重要であり、リファンピシンは組織や膿瘍に良く浸透し、表在性ブドウ球菌感染症の治療に一般的に使用されますが、耐性を迅速に誘導するため、他の薬剤と併用する必要があります。この研究の追加の発見として、相乗効果はMGOによるものではなく、MGOを添加した合成蜂蜜はリファンピシンとの相乗効果を示さなかったことが挙げられます。
作用機序がよく特徴付けられた抗菌剤の作用に対する蜂蜜の影響を理解することは、蜂蜜が細菌病原体にどのように影響するかの理解を深めることにもつながります。Liuら(2014年)は、相乗効果の解析を追加の抗生物質および異なるS. aureusおよびMRSA株に拡大しました。彼らは、蜂蜜によるタンパク質合成の抑制と抗生物質によるリボソームの阻害の相乗効果により、クリンダマイシンおよびゲンタマイシンに対する感受性の増加が生じる可能性があると示唆しました。一方、β-ラクタム系抗生物質との相乗効果は、両者による酸化ストレスの増加による可能性があります。S. aureusおよびMRSA株がオキサシリンと蜂蜜の組み合わせに同等に感受性を示したことから、相乗効果はPBP2Aのダウンレギュレーションによるものではないと考えられました。しかし、ある臨床MRSA分離株では、蜂蜜の存在下でクリンダマイシンやゲンタマイシンに対する感受性の増加が見られず、これはMRSAとS. aureusの蜂蜜に対する反応の違いが報告された初めてのケースとして注目されます。この株特異的な違いをトランスクリプトミクスやプロテオミクス解析で調査することは、将来の研究の興味深い方向性となるでしょう(Liuら、2014年)。
動物実験、症例報告、および臨床試験からの有効性の証拠
マヌカハニーを生産・販売する企業は高い倫理基準を推進し、感染症や創傷治癒の研究に動物モデルの使用を控えるよう促しています。しかし、マヌカハニーは特に馬の外科的または偶発的な創傷の治療に用いられ、良好な結果が報告されています(Dart et al., 2015; Bischofberger et al., 2016)。治癒しない創傷や潰瘍に対するハチミツ使用の症例報告では、従来の抗生物質が効果を示さなかった感染症の解消と著しい改善が見られています(Regulski, 2008; Smith et al., 2009)。しかし、これらや多くのin vitroおよびin vivoモデルでハチミツが問題となる創傷病原体を殺すという証拠があるにもかかわらず、マヌカハニーの堅牢な臨床データは不足しています。これは、ハチミツのような特徴的な物質に対する二重盲検プラセボ対照試験の技術的困難、倫理的配慮、臨床医の関心の欠如、自然製品と店頭販売に焦点を当てているハチミツ企業の費用対効果の問題など、さまざまな理由によります。これらは抗生物質耐性が現在の治療選択肢を侵食し続け、ハチミツの可能性を示す研究が医療従事者の注目を集めるにつれて変わる可能性があります。
ハチミツ研究におけるギャップと新たな機会
治療用ハチミツの理解は最近大きく進展しましたが、従来の抗生物質が効果を失い始めている場合でも、臨床医療での使用は依然として限られています。多様な病原体を殺し耐性を防ぐという点でハチミツの複雑さは最大の強みである一方で、多くの要因が相互に作用して活性に影響を与えるため、その研究は複雑です。私たちは、適切に登録された治療用マヌカハニーを用いたさらなる機構的研究、特に還元主義的でないシステム生物学的アプローチと詳細な化学的・微生物学的分析を組み合わせて、ハチミツが分子、細胞、集団レベルでどのように作用するか、異なる微生物株や種でどのように異なるか、そして宿主細胞がどのように反応するかを解明する研究を推奨します(表 2)。これらの研究から得られた情報は、治療に役立てられ、ハチミツを主流の医療に取り入れるために必要な臨床データを生み出すことができます。もはやすべてが失敗した場合にのみ使用される代替療法ではありません。
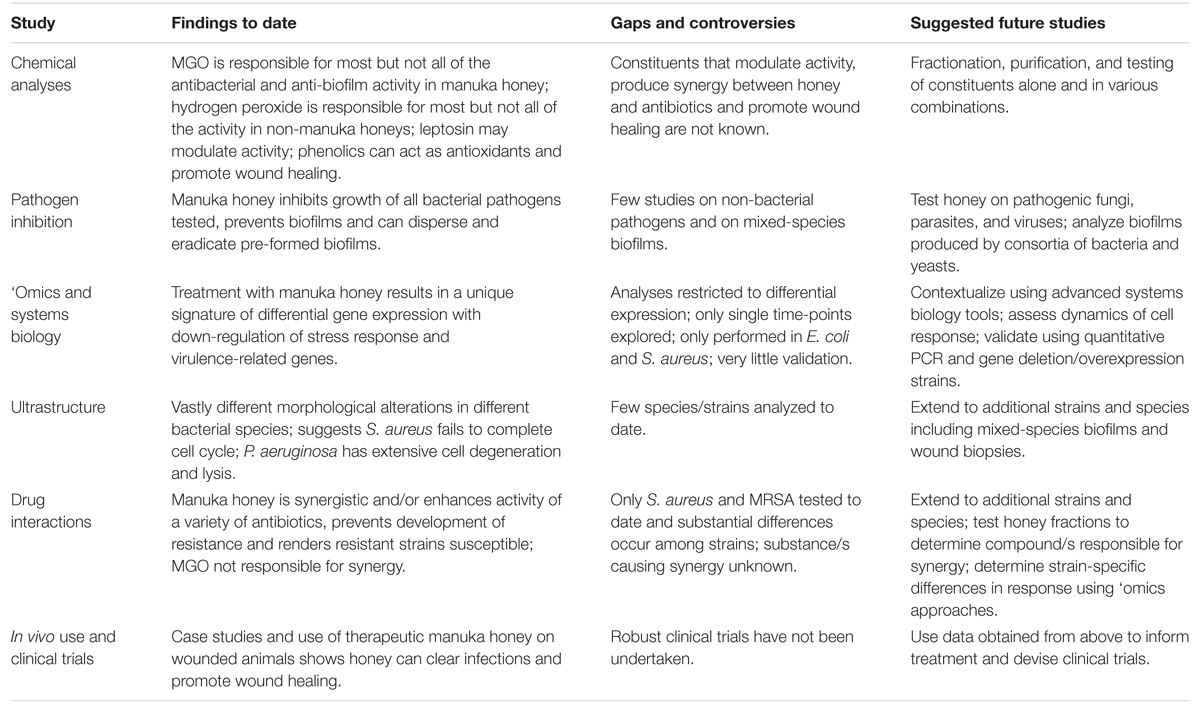
表2. マヌカハニーの研究:発見、ギャップ、今後の研究。
著者の貢献
このレビューはDC、SB、NNC、DB、PBによって執筆され、RSとEHによって批判的にレビューされました。
資金提供
NNCは農村産業研究開発公社-ハチプログラム(助成金PRJ-009186)から給与支援を受けています。
利益相反声明
DC、PB、EHはComvita NZ LimitedおよびCapilano Honey Limitedからのマヌカハニーの助成金および非財政的支援を報告しています。RSは医療用マヌカハニー(Medihoney)を取り扱うComvita NZ Limitedに雇用されています。
その他の著者は、本研究が潜在的な利益相反とみなされる商業的または財政的関係なしに実施されたことを宣言します。
略語
ESBL、拡張スペクトラムβ-ラクタマーゼ;MBC、最小殺菌濃度;MGO、メチルグリオキサール;MIC、最小阻止濃度;MRSA、メチシリン耐性Staphylococcus aureus;MRSE、メチシリン耐性Staphylococcus epidermis;NPA、非過酸化物活性;VRE、バンコマイシン耐性Enterococcus。
参考文献
Adams, C. J., Boult, C. H., Deadman, B. J., Farr, J. M., Grainger, M. N. C., Manley-Harris, M., et al. (2008). HPLCによる分離とニュージーランドのマヌカ(Leptospermum scoparium)蜂蜜の生物活性成分の特性評価。Carbohydr. Res. 343, 651–659. doi: 10.1016/j.carres.2007.12.011
PubMed要約 | CrossRef全文 |
Adams, C. J., Manley-Harris, M., and Molan, P. C. (2009). ニュージーランドのマヌカ(Leptospermum scoparium)蜂蜜におけるメチルグリオキサールの起源。Carbohydr. Res. 344, 1050–1053. doi: 10.1016/j.carres.2009.03.020
PubMed要約 | CrossRef全文 |
Al Somal, N., Coley, K. E., Molan, P. C., and Hancock, B. M. (1994). Helicobacter pyloriに対するマヌカハニーの抗菌活性の感受性。J. R. Soc. Med. 87, 9–12.
PubMed要約 |
Allen, K., Molan, P., and Reid, G. (1991). ニュージーランドのいくつかの蜂蜜の抗菌活性に関する調査。J. Pharm. Pharmacol.43, 817–822. doi: 10.1111/j.2042-7158.1991.tb03186.x
Allen, K. L., and Molan, P. C. (1997). 乳房炎を引き起こす細菌のハチミツの抗菌活性に対する感受性。N. Z. J. Agric. Res. 40, 537–540. doi: 10.1080/00288233.1997.9513276
Anthimidou, E., and Mossialos, D. (2012). ギリシャおよびキプロスのハチミツのStaphylococcus aureusおよびPseudomonas aeruginosaに対する抗菌活性のManukaハニーとの比較。J. Med. Food 16, 42–47. doi: 10.1089/jmf.2012.0042
PubMed要約 | CrossRef全文 |
Balan, P., Mal, G., Das, S., and Singh, H. (2016). クルクミン、Manukaハニーおよびホエイタンパク質の相乗的および加算的抗菌活性。J. Food Biochem. doi: 10.1111/jfbc.12249
Bassam, Z., Zohra, B. I., and Saada, A.-A. (1997). ハチミツのLeishmania寄生虫に対する効果:in vitro研究。Trop. Doctor 27, 36–38.
PubMed要約 |
Biglari, B., Moghaddam, A., Santos, K., Blaser, G., Büchler, A., Jansen, G., 他 (2013). ハニー(Medihoney)を用いた専門的創傷ケアに関する多施設前向き観察研究。Int. Wound J. 10, 252–259. doi: 10.1111/j.1742-481X.2012.00970.x
PubMed要約 | CrossRef全文 |
Bischofberger, A., Dart, C., Horadagoda, N., Perkins, N., Jeffcott, L., Little, C., 他 (2016). 馬の遠位肢の汚染された全層皮膚創傷におけるManukaハニーゲルのトランスフォーミング成長因子β1およびβ3濃度、細菌数および組織形態への影響。Aust. Vet. J. 94, 27–34. doi: 10.1111/avj.12405
PubMed要約 | CrossRef全文 |
Blair, S., Cokcetin, N., Harry, E., and Carter, D. (2009). 医療用グレードのLeptospermum蜂蜜の特異な抗菌活性:抗菌スペクトル、耐性およびトランスクリプトーム解析。Eur. J. Clin. Microbiol. Infect. Dis. 28, 1199–1208. doi: 10.1007/s10096-009-0763-z
PubMed要約 | CrossRef全文 |
Brady, N., Molan, P., and Harfoot, C. (1996). Manuka蜂蜜および他の蜂蜜の抗菌活性に対する皮膚糸状菌の感受性。Pharm. Pharmacol. Commun. 2, 471–473.
Carnwath, R., Graham, E. M., Reynolds, K., and Pollock, P. J. (2014). 馬の一般的な創傷細菌分離株に対する蜂蜜の抗菌活性。Vet. J. 199, 110–114. doi: 10.1016/j.tvjl.2013.07.003
PubMed要約 | CrossRef全文 |
Chen, C., Campbell, L., Blair, S. E., and Carter, D. A. (2012). 加熱処理が蜂蜜の抗菌特性に与える影響。Front. Microbiol. 3:265. doi: 10.3389/fmicb.2012.00265
Cooper, R., Jenkins, L., Henriques, A., Duggan, R., and Burton, N. (2010). 医療用グレードのManuka蜂蜜に対する細菌の耐性の不在。Eur. J. Clin. Microbiol. Infect. Dis. 29, 1237–1241. doi: 10.1007/s10096-010-0992-1
PubMed要約 | CrossRef全文 |
Cooper, R. A., Halas, E., and Molan, P. C. (2002a). 感染した火傷から分離されたPseudomonas aeruginosa株の抑制における蜂蜜の有効性。J. Burn Care Rehabil. 23, 366–370. doi: 10.1097/00004630-200211000-00002
PubMed要約 | CrossRef全文 |
Cooper, R. A., and Molan, P. C. (1999). Pseudomonas感染管理における蜂蜜の防腐剤としての使用。 J. Wound Care 8, 161–164. doi: 10.12968/jowc.1999.8.4.25867
PubMed要約 | CrossRef全文 |
Cooper, R. A., Molan, P. C., and Harding, K. G. (1999). 感染創傷からのStaphylococcus aureus株に対する蜂蜜の抗菌活性。 J. R. Soc. Med. 92, 283–285.
PubMed要約 |
Cooper, R. A., Molan, P. C., and Harding, K. G. (2002b). 創傷から分離された臨床的に重要なグラム陽性球菌の蜂蜜に対する感受性。 J. Appl. Microbiol. 93, 857–863. doi: 10.1046/j.1365-2672.2002.01761.x
PubMed要約 | CrossRef全文 |
Cooper, R. A., Wigley, P., and Burton, N. F. (2000). 多剤耐性株のBurkholderia cepaciaに対する蜂蜜の感受性。 Lett. Appl. Microbiol. 31, 20–24. doi: 10.1046/j.1472-765x.2000.00756.x
PubMed要約 | CrossRef全文 |
Dart, A., Bischofberger, A., Dart, C., and Jeffcott, L. (2015). マヌカハニーを用いた二次治癒馬の創傷治癒に関する研究レビュー:現在の推奨事項と将来の応用。 Equine Vet. Educ. 27, 658–664. doi: 10.1111/eve.12379
Estevinho, L., Pereira, A. P., Moreira, L., Dias, L. G., and Pereira, E. (2008). ポルトガル北東部の蜂蜜のフェノール化合物抽出物の抗酸化および抗菌効果。 Food Chem. Toxicol. 46, 3774–3779. doi: 10.1016/j.fct.2008.09.062
PubMed要約 | CrossRef全文 |
French, V. M., Cooper, R. A., and Molan, P. C. (2005). コアグラーゼ陰性ブドウ球菌に対する蜂蜜の抗菌活性。J. Antimicrobial Chemother. 56, 228–231. doi: 10.1093/jac/dki193
PubMed要約 | CrossRef全文 |
George, N. M., and Cutting, K. F. (2007). 抗菌蜂蜜(Medihoney):MRSA、VRE、およびPseudomonas aeruginosaを含む多剤耐性グラム陰性菌の臨床分離株に対するin vitro活性。Wounds 19:231.
PubMed要約 |
Halstead, F. D., Webber, M. A., Rauf, M., Burt, R., Dryden, M., and Oppenheim, B. A. (2016). エンジニアードハニー、医療用グレードの蜂蜜、および抗菌創傷被覆材のin vitro活性:バイオフィルム産生臨床細菌分離株に対する効果。J. Wound Care 25, 93–102. doi: 10.12968/jowc.2016.25.2.93
PubMed要約 | CrossRef全文 |
Hammond, E. N., and Donkor, E. S. (2013). Manuka蜂蜜のClostridium difficileに対する抗菌効果。BMC Res. 6:188. doi: 10.1186/1756-0500-6-188
PubMed要約 | CrossRef全文 |
Henriques, A. F., Jenkins, R. E., Burton, N. F., and Cooper, R. A. (2010). Manuka蜂蜜のStaphylococcus aureusに対する細胞内効果。Eur. J. Clin. Microbiol. Infect. Dis. 29, 45–50. doi: 10.1007/s10096-009-0817-2
PubMed要約 | CrossRef全文 |
Henriques, A. F., Jenkins, R. E., Burton, N. F., and Cooper, R. A. (2011). Manuka蜂蜜がPseudomonas aeruginosaの構造に与える影響。Eur. J. Clin. Microbiol. Infect. Dis. 30, 167–171. doi: 10.1007/s10096-010-1065-1
PubMed要約 | CrossRef全文 |
Hudson, N. J., Dalrymple, B. P., and Reverter, A. (2012). 発現差を超えて:原因となる変異と効果分子の探索。BMC Genomics 13:356. doi: 10.1186/1471-2164-13-356
PubMed要約 | CrossRef全文 |
Irish, J., Blair, S., and Carter, D. (2011). オーストラリアの植物由来のハチミツの抗菌活性。PLoS ONE 6:e18229. doi: 10.1371/journal.pone.0018229
PubMed要約 | CrossRef全文 |
Irish, J., Carter, D. A., Shokohi, T., and Blair, S. E. (2006). ハチミツはCandida種に対して抗真菌効果を持つ。Med. Mycol.44, 289–291. doi: 10.1080/13693780600931986
PubMed要約 | CrossRef全文 |
Jenkins, R., Burton, N., and Cooper, R. (2011). メチシリン耐性Staphylococcus aureusにおけるユニバーサルストレスプロテインAの発現に対するManukaハニーの影響。Int. J. Antimicrob. Agents 37, 373–376. doi: 10.1016/j.ijantimicag.2010.11.036
PubMed要約 | CrossRef全文 |
Jenkins, R., Burton, N., and Cooper, R. (2013). メチシリン耐性Staphylococcus aureus(MRSA)をin vitroでManukaハニーに曝露した際のプロテオミクスおよびゲノム解析により、病原性マーカーのダウンレギュレーションが示された。J. Antimicrobial Chemother. 69, 603–615. doi: 10.1093/jac/dkt430
PubMed 抄録 | CrossRef 全文 |
Jenkins, R., and Cooper, R. (2012). マヌカハニーを用いた創傷病原体に対する抗生物質活性のin vitro改善。PLoS ONE 7:e45600. doi: 10.1371/journal.pone.0045600
PubMed 抄録 | CrossRef 全文 |
Jenkins, R., Roberts, A., and Brown, H. L. (2015a). マヌカハニーの抗菌効果に関する機序的洞察。Res. Rep. Biol. 6, 215–224. doi: 10.2147/RRB.S75754
Jenkins, R., Wootton, M., Howe, R., and Cooper, R. (2015b). 嚢胞性線維症患者から得られた臨床分離株のマヌカハニーに対する感受性の実証。Arch. Microbiol. 197, 597–601. doi: 10.1007/s00203-015-1091-6
PubMed 抄録 | CrossRef 全文 |
Kalapos, M. P. (2008). フリーラジカルとメチルグリオキサールの連携。Chem. Biol. Interact. 171, 251–271. doi: 10.1016/j.cbi.2007.11.009
PubMed 抄録 | CrossRef 全文 |
Kato, Y., Umeda, N., Maeda, A., Matsumoto, D., Kitamoto, N., and Kikuzaki, H. (2012). 新規グリコシド、レプトシンの同定:マヌカハニーの化学的マーカーとして。J. Agric. Food Chem. 60, 3418–3423. doi: 10.1021/jf300068w
PubMed 抄録 | CrossRef 全文 |
Kilty, S. J., Duval, M., Chan, F. T., Ferris, W., and Slinger, R. (2011). メチルグリオキサール:(マヌカハニーの活性成分)in vitroにおける細菌バイオフィルムに対する活性。Int. Forum Allergy Rhinol. 1, 348–350. doi: 10.1002/alr.20073
PubMed要約 | CrossRef全文 |
Kronda, J. M., Cooper, R. A., Maddocks, S. E. (2013). ManukaハニーはPseudomonas aeruginosaのサイドロフォア産生を抑制する。J. Appl. Microbiol. 115, 86–90. doi: 10.1111/jam.12222
PubMed要約 | CrossRef全文 |
Kwakman, P. H., te Velde, A. A., de Boer, L., Vandenbroucke-Grauls, C. M., Zaat, S. A. (2011). 2つの主要な医療用ハニーは異なる殺菌メカニズムを持つ。PLoS ONE 6:e17709. doi: 10.1371/journal.pone.0017709
PubMed要約 | CrossRef全文 |
Lee, J.-H., Park, J.-H., Kim, J.-A., Neupane, G. P., Cho, M. H., Lee, C.-S., 他 (2011). 低濃度のハニーはEscherichia coli O157:H7のバイオフィルム形成、クオラムセンシング、毒性を低減する。Biofouling 27, 1095–1104. doi: 10.1080/08927014.2011.633704
PubMed要約 | CrossRef全文 |
Lin, S. M., Molan, P. C., Cursons, R. T. (2011). Manukaハニーの抗菌効果に対する消化管病原体の制御されたin vitro感受性。Eur. J. Clin. Microbiol. Infect. Dis. 30, 569–574. doi: 10.1007/s10096-010-1121-x
PubMed要約 | CrossRef全文 |
Liu, M., Lu, J., Müller, P., Turnbull, L., Burke, C. M., Schlothauer, R. C., 他 (2014). 抗生物質特異的なStaphylococcus aureusの抗菌剤とManukaハニー併用治療に対する反応の違い。Front. Microbiol. 5:779. doi: 10.3389/fmicb.2014.00779
PubMed 抄録 | CrossRef 全文 |
Lu, J., Carter, D. A., Turnbull, L., Rosendale, D., Hedderley, D., Stephens, J., et al. (2013). ニュージーランドのカヌカ、Manuka、クローバーのハチミツが細菌の成長動態と細胞形態に与える影響は種によって異なる。PLoS ONE 8:e55898. doi: 10.1371/journal.pone.0055898
PubMed 抄録 | CrossRef 全文 |
Lu, J., Turnbull, L., Burke, C. M., Liu, M., Carter, D. A., Schlothauer, R. C., et al. (2014). Manuka タイプのハチミツは異なるバイオフィルム形成能力を持つStaphylococcus aureus株が作るバイオフィルムを根絶できる。PeerJ 2:e326. doi: 10.7717/peerj.326
PubMed 抄録 | CrossRef 全文 |
Maddocks, S. E., Jenkins, R. E., Rowlands, R. S., Purdy, K. J., and Cooper, R. A. (2013). Manuka ハチミツは医療的に重要な創傷細菌の付着および侵入をin vitroで抑制する。Fut. Microbiol. 8, 1523–1536. doi: 10.2217/fmb.13.126
PubMed 抄録 | CrossRef 全文 |
Maddocks, S. E., Lopez, M. S., Rowlands, R. S., and Cooper, R. A. (2012). Manuka ハチミツはStreptococcus pyogenesのバイオフィルム形成を抑制し、2つのフィブロネクチン結合タンパク質の発現を低下させる。Microbiology 158, 781–790. doi: 10.1099/mic.0.053959-0
PubMed 抄録 | CrossRef 全文 |
Majtan, J. (2014). ハチミツ:創傷治癒における免疫調節因子。Wound Repair Regenerat. 22, 187–192. doi: 10.1111/wrr.12117
PubMed 抄録 | CrossRef 全文 |
Majtan, J., Bohova, J., Horniackova, M., Klaudiny, J., and Majtan, V. (2014a). 創傷病原体Proteus mirabilisおよびEnterobacter cloacaeに対する蜂蜜の抗バイオフィルム効果。Phytother. Res. 28, 69–75. doi: 10.1002/ptr.4957
PubMed 抄録 | CrossRef 全文 |
Majtan, J., Bohova, J., Prochazka, E., and Klaudiny, J. (2014b). メチルグリオキサールはグルコースオキシダーゼの阻害を通じてマヌカ蜂蜜中の過酸化水素蓄積に影響を与える可能性がある。J. Med. Food 17, 290–293. doi: 10.1089/jmf.2012.0201
PubMed 抄録 | CrossRef 全文 |
Majtan, J., Klaudiny, J., Bohova, J., Kohutova, L., Dzurova, M., Sediva, M., et al. (2012). マヌカ蜂蜜中の重要なミツバチタンパク質成分に対するメチルグリオキサール誘発修飾:治療的意義の可能性。Fitoterapia 83, 671–677. doi: 10.1016/j.fitote.2012.02.002
PubMed 抄録 | CrossRef 全文 |
Majtan, J., Majtanova, L., Bohova, J., and Majtan, V. (2011). 癌患者から分離された多剤耐性Stenotrophomonas maltophilia株の根絶における強力な抗菌剤としてのハニーデュー蜂蜜。Phytother. Res. 25, 584–587. doi: 10.1002/ptr.3304
PubMed 抄録 | CrossRef 全文 |
Malone, L. A., Gatehouse, H. S., and Tregidga, E. L. (2001). ミツバチの寄生虫であるNosema apis(Microsporidia: Nosematidae)に対する時間、温度、および蜂蜜の影響。J. Invertebrate Pathol. 77, 258–268. doi: 10.1006/jipa.2001.5028
PubMed要約 | CrossRef全文 |
Mavric, E., Wittmann, S., Barth, G., and Henle, T. (2008). ニュージーランド産Manuka(Leptospermum scoparium)はちみつの主要な抗菌成分としてのメチルグリオキサールの同定と定量。 Mol. Nutrit. Food Res. 52, 483–489. doi: 10.1002/mnfr.200700282
PubMed要約 | CrossRef全文 |
Mullai, V., and Menon, T. (2007). 臨床および環境由来のPseudomonas aeruginosa分離株に対する異なる種類のはちみつの殺菌活性。 J. Alternat. Complement. Med. 13, 439–442. doi: 10.1089/acm.2007.6366
PubMed要約 | CrossRef全文 |
Müller, P., Alber, D. G., Turnbull, L., Schlothauer, R. C., Carter, D. A., Whitchurch, C. B., et al. (2013). メディハニーとリファンピシンのメチシリン耐性Staphylococcus aureus(MRSA)に対する相乗効果。 PLoS ONE 8:e57679. doi: 10.1371/journal.pone.0057679
PubMed要約 | CrossRef全文 |
Mundo, M. A., Padilla-Zakour, O. I., and Worobo, R. W. (2004). 選択された生はちみつによる食品由来病原菌および食品腐敗微生物の増殖抑制。 Int. J. Food Microbiol. 97, 1–8. doi: 10.1016/j.ijfoodmicro.2004.03.025
PubMed要約 | CrossRef全文 |
Nilforoushzadeh, M. A., Jaffary, F., Moradi, S., Derakhshan, R., Haftbaradaran, E. (2007). 皮膚リーシュマニア症治療における局所ハチミツ塗布とグルカンタイムの局所注射の効果。BMC Complement Altern. Med. 7:1. doi: 10.1186/1472-6882-7-1
Norton, A. M., McKenzie, L. N., Brooks, P. R., Pappalardo, L. J. (2015). 高速液体クロマトグラフィーによるオーストラリア産Leptospermumの蜜に含まれるジヒドロキシアセトンの定量。J. Agric. Food Chem. 63, 6513–6517. doi: 10.1021/acs.jafc.5b01930
Okhiria, O., Henriques, A., Burton, N., Peters, A., Cooper, R. (2009). ハチミツはPseudomonas aeruginosaのバイオフィルムを時間および用量依存的に調節する。J. ApiProduct. ApiMedical Sci. 1, 6–10. doi: 10.3896/IBRA.4.01.1.03
Osato, M. S., Reddy, S. G., Graham, D. Y. (1999). ハチミツの浸透圧効果がHelicobacter pyloriの成長と生存に及ぼす影響。Dig. Dis. Sci. 44, 462–464. doi: 10.1023/A:1026676517213
Packer, J. M., Irish, J., Herbert, B. R., Hill, C., Padula, M., Blair, S. E., 他 (2012). ManukaハニーのStaphylococcus aureusプロテオームに対する特異的な非過酸化物抗菌効果。Int. J. Antimicrob. Agents 40, 43–50. doi: 10.1016/j.ijantimicag.2012.03.012
Roberts, A. E., Maddocks, S. E., and Cooper, R. A. (2012). ManukaハニーはPseudomonas aeruginosaに対して殺菌的であり、oprFおよびalgDの発現に差異をもたらす。Microbiology 158, 3005–3013. doi: 10.1099/mic.0.062794-0
PubMed要約 | CrossRef全文 |
Roberts, A. E. L., Maddocks, S. E., and Cooper, R. A. (2015). Manukaハニーは鞭毛関連遺伝子の抑制によりPseudomonas aeruginosaの運動性を低下させる。J. Antimicrob. Chemother. 70, 716–725. doi: 10.1093/jac/dku448
PubMed要約 | CrossRef全文 |
Sajid, M., and Azim, M. K. (2012). 天然ハニーの線虫殺傷活性の特性評価。J. Agric. Food Chem. 60, 7428–7434. doi: 10.1021/jf301653n
PubMed要約 | CrossRef全文 |
Shahzad, A., and Cohrs, R. J. (2012). 水痘帯状疱疹ウイルス(VZV)に対するハニーのin vitro抗ウイルス活性:帯状疱疹の潜在的治療法のためのトランスレーショナルメディシン研究。Transl. Biomed. 3:2.
PubMed要約 |
Sherlock, O., Dolan, A., Athman, R., Power, A., Gethin, G., Cowman, S., 他 (2010). チリ産UlmoハニーとManukaハニーのメチシリン耐性Staphylococcus aureus、Escherichia coli、およびPseudomonas aeruginosaに対する抗菌活性の比較。BMC Complement Altern. Med. 10:47. doi: 10.1186/1472-6882-10-47
PubMed要約 | CrossRef全文 |
Silva-Rocha, R., and de Lorenzo, V. (2010). 原核生物の調節ネットワークにおけるノイズとロバスト性。Annu. Rev. Microbiol. 64, 257–275. doi: 10.1146/annurev.micro.091208.073229
PubMed要約 | CrossRef全文 |
Smith, T., Hanft, J. R., Legel, K. (2009). 難治性静脈性脚創傷に対する局所的Leptospermumハニーの予備的症例シリーズ。 Adv. Skin Wound Care 22, 68–71. doi: 10.1097/01.ASW.0000345283.05532.9a
PubMed要約 | CrossRef全文 |
Soffer, A. (1976). チワワ犬とラエトリル、キレート療法、コロラド州ボルダー産ハニー [editorial]。 Arch. Intern. Med.136, 865–866. doi: 10.1001/archinte.136.8.865
PubMed要約 | CrossRef全文 |
Sojka, M., Valachova, I., Bucekova, M., Majtan, J. (2016). 多種創傷バイオフィルムに対するハニーおよびミツバチ由来デフェンシン-1の抗バイオフィルム効果。 J. Med. Microbiol. doi: 10.1099/jmm.0.000227 [Epub ahead of print].
PubMed要約 | CrossRef全文 |
Stephens, J. M., Schlothauer, R. C., Morris, B. D., Yang, D., Fearnley, L., Greenwood, D. R., 他 (2010). ニュージーランド産ManukaおよびKanukaハニーのフェノール化合物とメチルグリオキサール。 Food Chem. 120, 78–86. doi: 10.1016/j.foodchem.2009.09.074
Tan, H. T., Rahman, R. A., Gan, S. H., Halim, A. S., Hassan, S. A., Sulaiman, S. A., 他 (2009). マレーシア産トゥアラングハニーの創傷および腸内微生物に対する抗菌特性のManukaハニーとの比較。 BMC Complement Altern. Med. 9:34. doi: 10.1186/1472-6882-9-34
PubMed要約 | CrossRef全文 |
Watanabe, K., Rahmasari, R., Matsunaga, A., Haruyama, T., and Kobayashi, N. (2014). ハニーのin vitroにおける抗インフルエンザウイルス効果:Manukaハニーの強力な高活性。Arch. Med. Res. 45, 359–365. doi: 10.1016/j.arcmed.2014.05.006
PubMed要約 | CrossRef全文 |
Wilkinson, J. M., and Cavanagh, H. M. A. (2005). 13種類のハニーのEscherichia coliおよびPseudomonas aeruginosaに対する抗菌活性。J. Med. Food 8, 100–103. doi: 10.1089/jmf.2005.8.100
PubMed要約 | CrossRef全文 |
Williams, S., King, J., Revell, M., Manley-Harris, M., Balks, M., Janusch, F., 他 (2014). ニュージーランドのManuka (Leptospermum scoparium)の花蜜中ジヒドロキシアセトン含有量の地域的、年次的、個体差。J. Agric. Food Chem. 62, 10332–10340. doi: 10.1021/jf5045958
PubMed要約 | CrossRef全文 |
Willix, D. J., Molan, P. C., and Harfoot, C. G. (1992). 傷に感染する細菌種のManukaハニーおよび他のハニーの抗菌活性に対する感受性の比較。J. Appl. Bacteriol. 73, 388–394. doi: 10.1111/j.1365-2672.1992.tb04993.x
PubMed要約 | CrossRef全文 |
Windsor, S., Pappalardo, M., Brooks, P., Williams, S., and Manley-Harris, M. (2012). オーストラリア産Leptospermumハニーに適用したジヒドロキシアセトンとメチルグリオキサールの便利な新分析法。J. Pharmacogn. Phytother. 4, 6–11.
治療用Manukaハニー:もはや代替ではない - https://www.frontiersin.org/articles/10.3389/fmicb.2016.00569/full
コメントを残す